Article Text

Statistics from Altmetric.com
Introduction
Advances in acute stroke treatment and the widespread establishment of dedicated stroke units have resulted in an increase in poststroke survival and life expectancy. However, stroke remains a leading cause of long-term disability worldwide, making the improvement of poststroke outcomes a chief healthcare goal for many countries. The current strategies strive to reduce the initial injury by acutely implementing thrombolytic and/or endovascular interventions, to better understand the major determinants that influence the stroke recovery and to search for innovative, effective and accessible recovery and rehabilitation modalities that can mitigate various poststroke deficits and enhance the quality of life. These approaches require a collaboration and integration of fundamental and clinical science research to more efficiently translate benchwork results into therapeutic bedside interventions. Due to a variety of stroke research advances in both the basic and clinical sciences over the last few years, especially in 2016, the field of stroke recovery and rehabilitation has celebrated many hopes and progresses. Our goal was to explore these studies and better identify, understand and integrate key findings for the purpose of identifying new targets that could be translated into clinically rewarding therapeutic interventions in future.
We manually searched professional journals with an average 5-year impact factor >3 (from 2012 to 2016) that were known to publish manuscript with topics in stroke recovery and rehabilitation. We aimed to selectively highlight relevant basic and clinical science stroke recovery research published between December 2015 and December 2016 in these journals. Certain selection biases cannot be completely ruled out and omissions are possible. The list of journals are Science, Nature, Nature Neuroscience, Neuron, Proceedings of the National Academy of Sciences of the United States of America, Neurobiology of Disease, Scientific Report, PLOS ONE, Acta Neuropathologica, Journal of Neuroscience, Annals of Neurology, Neurology, JAMA Neurology, Stroke, The Lancet, Lancet of Neurology, JAMA, Brain, Brain Stimulation, Stem Cells, Cell Death and Differentiation, Neurorehabilitation and Neural Repair, Journal of Cerebral Blood Flow and Metabolism and New England Journal of Medicine.
Basic science
Blood–brain barrier
Amidst the frustration of finding effective neuroprotective agents, several researchers attempted to further investigate the blood–brainbarrier (BBB)’s role in protecting against ischaemic injury and to find new methods of repairing ischaemia-induced BBB dysfunction. Gurnik S et al studied the specific role of angiotensin-2 (Ang-2) in stroke-induced BBB dysfunction.1 In this study, they investigated the influence of Ang-2 on BBB permeability in wild-type and gain-of-function (GOF) mice. They found that the Ang-2 GOF mice, which express Ang-2 in an inducible, endothelial-specific manner, exhibited decreased pericyte coverage of vessels and increased caveolae-like vesicles in endothelial cells (ECs), as well as increased permeability of BBB. In addition, after wild-type and Ang-2 GOF mice were subjected to either permanent middle cerebral artery occlusion (MCAO) or only 60 min MCAO, the Ang-2 GOF mice showed larger infarcts and higher BBB permeability. To further demonstrate the concept, they used the vascular endothelial protein tyrosine phosphatase inhibitor AKB-9785 to block the Ang-2 expression which indeed ameliorated the deficits. Therefore, Ang-2 could be a potential therapeutic target for protecting BBB integrity during ischaemic strokes that need to be tested in human studies.
Another potential player, the matrix metalloproteinase-9 (MMP-9), is believed to mediate ischaemia-induced BBB disruption by degrading endothelial junction proteins and basal lamina in the initial phase after ischaemia. Shi et al, however, elucidated that while the late-onset (3–6 hours post ischaemia–reperfusion injury) BBB leakage of large molecules did depend on MMP-9, the early and subtle BBB leakage within 30–60 min after the ischaemia–reperfusion injury was independent of MMP-9 activities.2 This is important to distinguish, for this early, subtle increase in BBB permeability postischaemia may be partially reversible. Moreover, the authors found that the transgenic mice with EC-specific overexpression of constitutively active mutant actin-depolymerising factor (ADF) could potentially attenuate MMP-9-dependent delayed BBB injury caused after 60 min MCAO. This result was supported by evidence from neurobehavioural assessment as well as histological characterisation. Because of the potential reversibility of the early, subtle increase in BBB permeability after ischaemia, ADF would be an attractive therapeutic target for protecting the BBB and for reducing neuronal injury.
Efforts were also made to better understand the endogenous system and its role in restoring damaged BBB after injury. Lou et al investigated how juxtavascular microglial cells help to amend injured BBB.3 After they used 2-photon focused laser excitation to trigger capillary damage in small focal brain lesions, the authors found that the microglial cells in the perivascular region were attracted to the site of capillary lesions, where they formed a dense plexiform aggregate. Furthermore, photoablation of these microglial cells abolished this rapid closure of the BBB’s leakage. Since microglia are the only cells that expressed detectable levels of the purinergic receptor P2Y12 (P2RY12), it was not surprising that the P2RY12-deficient mouse failed to close the laser-induced BBB opening. Therefore, juxtavascular microglia are likely involved with repairing the damaged BBB via the P2RY12 receptor-mediated pathways.
Neuron apoptosis and autophagy
Apoptosis and autophagy are other target areas that are being extensively investigated in stroke recovery research. Peng et al 4 provided evidence that forkhead box O3a (FOXO3a) is a key transcription factor involved in neuronal apoptosis and showed that histone deacetylase 2 (HDAC2) selectively regulated FOXO3a-mediated gene transcription during oxidative stress-induced neuronal cell death. HDAC2 is an enzyme that belongs to the histone deacetylase family in humans. Knocking down HDAC2 can thus protect neurons from H2O2-induced apoptosis. A different group, Baik et al found that the peptidyl-prolyl cis-trans isomerase NIMA-interacting 1 (Pin1) contributed to neuronal apoptosis under ischaemic conditions via Notch signalling. Pin1, when bound, can activate Notch homologue 1 (Notch1) and stabilise the Notch intracellular domain. High expression of Pin 1 could promote cell death by accumulating the Notch intracellular domain.5
Autophagy, a lysosomal pathway for the recycling of organelles and long-lived proteins, is another type of programmed cell death. With regard to stroke, several vascular risk factors can increase brain cell injury via autophagy, like hyperhomocysteinaemia.6 Autophagy is a double-edged sword. An imbalance or excessive activation of autophagy promotes cellular pathology and ultimately leads to ischaemic cell death; therefore, suppression of autophagy by bexarotene7 or ganglioside GM18 can decrease autophagical cell death, reduces the infarct volume and even prevents secondary neurodegenerative injury. Autophagical cell death may not indeed be caused by excessive autophagy rather by the inhibition of autophagy flux, which is the degradation activity. In a certain stage of brain injury, overactivation of autophagic pathways could act as a protective mechanism.9
Role of microRNA in angiogenesis
Angiogenesis in perilesional areas has drawn significant attention throughout previous years of neurological research. In the past, microRNA and small non-coding RNA molecules are important in regulating post-transcriptional gene expression in the brain. Many miRNAs are highly expressed in the vasculature and serve critical modulators of vascular cell function. Recently, several studies demonstrated that a variety of miRNAs are involved in regulating angiogenesis after stroke. For example, miR-107 was significantly upregulated after permanent MCAO in rats, and inhibition of miR-107 resulted in reduced capillary density in the ischaemic boundary zone.10 In addition, they found that both ECs and astrocytes secreted miR-107 when exposed to hypoxia; the upregulated miR-107 acted mainly on ECs to promote tube formation and migration. Furthermore, by downregulating Dicer 1, a gene that encodes an enzyme needed for processing miRNA precursors, miR-107 can suppress the mRNA of vascular endothelial growth factor (VEGF) 164 and VEGF165. This upregulation of VGEF eventually leads to angiogenesis after stroke, signifying that miR-107 may be used as a novel tool for promoting stroke recovery.
Another study from Caballero-Garrido et al demonstrated that intravenous injections of a specific miR-155 inhibitor at 48 hours after distal MCAO (dMCAO) in mice could reduce infarct volume and improve the animal’s neuron function recovery; mechanistically, the inhibition of miR-155 resulted in increased endothelial nitric oxide production and reduction of vascular inflammation.11 The neuronal functional recovery was assessed by the bilateral asymmetry/adhesive removal test and Cat-Walk automated gait analysis system. Likewise, Chen et al showed that miR-126 contributes to human umbilical cord blood cell (HUCBC)-induced neurorestorative effects in type 2 diabetes mellitus (T2DM) mice.12 HUCBCs were used to treat a group of T2DM mice at 3 days after dMCAO and they promoted miR-126 expression, which subsequently enhanced axon and myelin density in the ischaemic brain.
MicroRNAs, in addition to their function in regulating angiogenesis described earlier, are also believed to act as regulators of synaptic signalling and are closely related to neuronal apoptosis. Using Drosophila models, Verma et al 13 showed that genetic deletion of Drosophila miR-1000 led to elevated apoptosis in the brain as a result of glutamatergic excitotoxicity; miR-1000 regulated the expression of the vesicular glutamate transporter (VGluT), which loads glutamate into synaptic vesicles. Similarly, when the mammalian miR-137 (seed-similar miRNA) was depleted in mouse cortical neuron cultures, a simultaneous increase in VGluT mRNA level was observed, accompanied by more caspase 3-positive cells. This indicates that this particular miRNA’s neuroprotective regulation of VGluT2 was conserved in mammals. Furthermore, VGluT protein levels were elevated when miR-137 was downexpressed in the dentate gyrus region of the hippocampus. Thus, miRNA-mediated regulation of glutamatergic activity works presynaptically to modulate synaptic transmission and to protect against excitotoxicity, which has been implicated in ischaemic brain injury.
Inflammasome and its contribution to inflammation and neural injury
Although mounting evidence has shown that inflammasome-mediated processes are involved with brain injury, the molecular mechanisms remain unclear. Inflammation after an acute stroke is driven by the proinflammatory cytokine interleukin-1 (IL-1) and is known to exacerbate the initial injury. The activity of IL-1 is regulated by multimolecular protein complexes called inflammasomes. Out of the plethora of inflammatory proteins involved in various disease conditions, two families were specifically implicated in ischaemic brain injury: the nucleotide-binding oligomerisation domain (NOD)-like receptor (NLR) family and the pyrin and haematopoietic interferon-inducible nuclear antigens (PYHIN) family. In a study by Denes et al,14 five kinds of knockout mice were used to prove that the NLR family CARD domain-containing protein 4 (NLRC4) and absence of melanoma 2 (AIM2) inflammasomes did indeed contribute to brain injury after ischaemia. NLRC4 belongs to the NLR family, and AIM2 belongs to the PYHIN family of inflammasomes. The authors found that both the NLRC4 knockout and AIM2 knockout mice were more resistant to ischaemia when compared with wild-type mice and therefore provided new insights on how the inflammatory response is regulated after acute ischaemia.
Although inflammasome-mediated processes in haematopoietic macrophages and dendritic cells have been extensively studied, little is known about their role in microglia. Burm et al 15 have discovered new and important cell type-specific differences in inflammasome-mediated responses by comparing inflammasome-mediated activation in primary microglia, bone marrow-derived macrophages and blood CD14+-derived macrophages from healthy adult rhesus macaques. For example, microglia and haematopoietic macrophages are generally endowed with similar inflammasome machinery, but microglia are less dependent than bone marrow-derived macrophages on inflammatory caspases for the processing and secretion of IL-1β. Understanding this variety in activation and responses can help to modulate innate immune responses in the brain.
Several other inflammasomes participating in the inflammation and injury were associated with ischaemic stroke. Kaushal et al 16 demonstrated that neuronal Nod-like receptor protein 1(NLRP1) inflammasome activation of caspase-1 modulates inflammatory IL-1β production and axonal degeneration-associated caspase-6 activation by analysing NLRP1 and caspase-1 knockout mice. Collectively, these findings provide mechanistic insight into how the inflammasomes help trigger neuroinflammation and axonal degeneration after stroke or brain injury.
Mitochondrial energy metabolism
Mitochondrion is one of the major organelles that maintain energy production in the cell environment and contribute to the development of apoptotic and necrotic cell death after stroke. In the central nervous system, neurons can transfer their damaged mitochondria to astrocytes for recycling. Hayakawa et al 17 investigated this exchange of mitochondria between neurons and astrocytes as a method of cell–cell signalling; he found that after transient focal ischaemia, astrocytes actively released functional mitochondria into adjacent neurons via a calcium-dependent mechanism involving CD38 and cyclic adenosine diphosphate(ADP) ribose signalling. The neighbouring neurons that incorporated those astrocyte-derived mitochondria became more resistant to the ischaemic stress. These findings suggest a new mitochondrial mechanism of neuroglial crosstalk that may contribute to endogenous neuroprotective and neurorecovery mechanism after stroke.
Chikka et al 18 investigated the interesting theory that immunological players originating outside of the nervous system can influence neuronal health. Specifically, mitochondrial dysfunction in intestinal cells can activate potentially neuroprotective immune pathways. The team exposed Caenorhabditis elegans to the mitotoxin rotenone stimulation as a model system of neurodegenerative disorders and they found that mitochondria in intestinal cells may protect neurons via a non-cell-autonomous fashion. The immune response pathway involved was P38MAPK/ATF-7 and its activation decreased oxidative damage by removing the dysfunctional mitochondria by mitochondrial autophagy and proved to be neuroprotective, the rotenone-induced dopaminergic neuron loss was suppressed when this pathway was enhanced. This study underpins the importance of the crosstalk between the gut and brain and further illustrates that poststroke inflammation can be mediated by the mitochondrion.
Another study also highlighted the central role of mitochondria in regulating neuronal function under diseased conditions. Wang et al 19 demonstrated mitochondrial localisation of TDP-43 (TAR DNA-binding protein 43) in neurons induced neuronal death after mitochondrial dysfunction and proposed that TDP-43 mitochondrial localisation can be a therapeutical target for neurodegeneration.
Cell-based approaches
Stem cell therapy has been extensively investigated as a potential regenerative strategy for patients who had a stroke with severe deficits. However, the majority of stem cells transplantation studies have not yet been successful in demonstrating good graft survival and long-term engraftment. Although mesenchymal stem cells (MSCs), with the features of easily isolated and amplified with low immunogenicity and of having a paracrine and immunomodulatory function, have been widely tested in preclinical animal studies, human clinical trials including the one from Steinberg et al 20 have been largely inconclusive. One possible explanation is low rate of neuronal cell replenishment.
Recently, neural stem cells (NSCs) have become cell candidates for transplantation in stroke models. Unfavourably, the endogenous host environment during the acute phase of stroke likely affects the survival rate of NSCs transplantation. Wakai et al 21 found that hypoxic preconditioning (5% hypoxia for 24 hours) significantly enhanced the viability of the NSCs exposed to haemoglobin and increased grafted cell survival in the intracerebral haemorrhage brain in mice. In addition, they found that more NSCs survived in the preconditioning group than the non-preconditioning group in the intracerebral haemorrhage if the transplanted NSCs underwent hypoxic preconditioning in vitro culture. The underlying mechanism is likely that hypoxic preconditioned NSCs exhibited more VEGF expression and enhanced peri-infarct region tissue protection, and subsequently improved functional recovery. This exciting result provided evidence that hypoxic preconditioning in NSCs could improve the efficacy of stem cell therapy.
Besides MSCs and NSCs, other stem cells, like multilineage-differentiating stress-enduring (Muse) cells, have been studied as well. Muse cell is unique stem cell population that is capable of self-renewal, stress tolerant and can differentiate into cells representative of all three germ layers from a single cell. Despite the fact that infarct size was not reduced, transplanted rats exhibited significant improvement in neurological and motor function at both day 70 and 84 as compared with the control group, Muse cells survived in the host brain for up to 84 days and integrated into the sensorimotor cortex, extending neurites even into the cervical spinal cord.22 These data suggest that Muse cells be a feasible and promising cell source for cell-based stroke therapy, and clinical study should be the next step. Although selecting effective cells for transplantation in stroke is critical, tracking the survival, differentiation and migration of transplanted cells into the brain are also a highly important and specialised process. Nicholls et al 23 proposed a novel, non-invasive method for visualising the distribution of transplanted NSCs and ECs. They used the used the paramagnetic chemical exchange saturation transfer agents (Eu-HPDO3A and Yb-HPDO3A) to label the human striatal NSC line STROC05 and the human cerebral microvascular ECs hCMEC/D3, respectively. Next, the implanted NSCs and ECs in vivo brains can be successfully visualised simultaneously and non-invasively by the magnetic resonance image checking, and accuracy was validated by histological analysis.
Brain–computer interface
While exogenous stem cells transplantation is promising as a viable strategy for brain repair, Capogrosso et al 24 chose to bypass attempts for repairing the brain injury and instead pursued an innovative technique that could restore behavioural function to the patients. The team recently developed a brain–spinal cord interface to decipher signals from the motor cortex that controls leg movement. In their study, they designed and implemented wireless control systems linking the online neural decoding of extension and flexion motor states with stimulation protocols; gait deficits after a complete spinal cord injury in a non-human primate were alleviated. These systems allowed the primate to behave freely without any restrictions or constraining tethered electronics. Although this paper is about brain–computer interface technology on a primate with a manipulated injury to its spinal cord, it clearly has a strong implication for use in stroke survivors with subcortical infarct severely damaging the corticospinal tract.
Clinical science
Birth of the stroke recovery and rehabilitation guideline
One of the most important milestones in 2016 was the release of the first ‘Guideline for adult stroke rehabilitation and recovery: a guideline for healthcare professional from American Heart Association/American Stroke Association’25by American Stroke Association. This guideline was endorsed by five professional rehabilitation societies—The American Academy of Physical Medicine and Rehabilitation, American Physical Therapy Association, American Occupational Therapy Association, American Society of Neurorehabilitation and American Congress of Rehabilitation Medicine. The guideline reflects collaborative work from many rehabilitation professionals. The total of 229 recommendations in the guideline address 41 recovery topics, and these recommendations were presented and organised into five primary categories (the rehabilitation programme, prevention and medical management of comorbidities, assessment, sensorimotor impairments and treatments/interventions, transitions in care and community rehabilitation). Now rehabilitation professionals have a universal document to guide their clinical practice.
Progress in motor recovery
Motor impairment is the most common deficit after stroke; therefore, reducing the motor deficit and improving outcomes were a chief goal for many clinical researchers in 2016. Recovery scientists continued to explore whether promoting one form of task-specific training or increasing the dose of rehabilitation therapy, which is likely underdosed in clinical practice, will lead to better motor improvement. The Interdisciplinary Comprehensive Arm Rehabilitation Evaluation (ICARE) Study26 had somewhat unexpected and disappointing results. It showed that a 12-month, structured, task-oriented upper extremity rehabilitation programme (an average of 30 hours of therapy) did not significantly improve motor function or recovery beyond either an equivalent (an average of 27 hours) or a lower dose of customary occupational therapy (an average of 11 hours). Based on the study, it can be said that a higher dose of rehabilitation therapy does not necessarily translate to better outcomes. This finding was further supported by another study by Lang et al. 27 28 In this single-centre, single-blind, parallel, randomised, controlled trial, 85 subjects with upper extremity paresis 6 months poststroke were randomised to one of four dose groups—3200 (100 repetitions/session), 6400 (200 repetitions/session), 9600 (100 repetitions/session) or individualised maximum repetitions (300 repetitions/session and sessions continuing until meeting prespecified stopping criteria), during 1 hour sessions, 4 days/week for 8 weeks. The dosing parameter manipulated was the amount of task-specific training, as indexed by the number of task repetitions above. The intervention was an individualised, progressive, task-specific upper limb training programme designed to improve upper limb functional motor capacity assessed by Action Research Arm Test. Overall, the treatment effect was minute and there was a lack of repetition–response relationship of task-specific training on motor functional recovery. After combining this with other findings from a previous study29, it is clear that a careful assessment of residual neurosubstrate is critical for stroke motor recovery.
According to several recent studies, virtual reality (VR) tool, like this innovative non-immersive VR using the Nintendo Wii gaming system (VRWii), as an add-on therapy to the conventional rehabilitation intervention is not better than a low-cost, simple recreational activity intervention in improving motor function, as measured by Wolf Motor Function Test (WMFT).29 30 Specifically, this four-nation single-blind randomised, controlled parallel 2-week study assigned 59 patients to the WRWii group and 62 patients to the recreational activity group: a 32.0% of reduction (or 14.0 s) was achieved in the WRWii group in performing WMFT versus a 28.2% (or 10.9 s) of reduction in the control group. The difference between the two groups was not statistically significant or clinically meaningful.
Patients who had a stroke will likely benefit from having multiple spasticity treatment options. Another botulinum toxin was recently demonstrated to be effective in reducing muscular tone and preventing spasticity in a multicentre placebo-controlled randomised clinical trial.31 Two hundred and forty-three subjects (with stroke or traumatic brain injury) were randomised (1:1:1 allocation) to receive a single injection session of abobotulinum toxin A 500 U or 1000 U or placebo into the most hypertonic muscles group among the elbow, wrist or finger flexors. At 4 weeks after injection, muscle tone assessed by modified Ashworth Spasticity(MAS) Scale decreased by 0.3 points (SD 0.6), 1.2 (SD 1.0) and 1.4 (SD 1.1) for placebo, 500 U and 1000 U group, respectively. Most importantly the difference in the changes of MAS between the both treatment group and placebo is statistically significant and clinically meaningful. Adverse events are minor, with mild muscle weakness being the most common one. One caution for interpretation of such data again is that botulinum toxin can reduce muscular tone, change the perception of limb deformity and it does not always lead to functional improvement. It is consistent with the recently updated guideline from American Academy of Neurology.32
Proportional recovery rule
The ‘Proportional recovery rule’ attracted renewed attentions in recent years.33 This phenomenon has been observed by several research groups, two in the USA34 35 and another in New Zealand36 where rehabilitation care systems are not same. Prabhakaran et al 35 first found that stroke patients with initial mild-to-moderate motor impairment showed an almost fixed proportional upper extremity motor recovery at 3 months. As also observed in a study by Bylow et al, 36 a patient who had a stroke with a positive Motor Evoked Potentials (MEPs, indicating a preserved corticomotor pathway) will recover approximately 70% of his/her maximally recoverable capacity at 12 weeks after stroke. The dosage of therapy had minimal effect on the degree of recovery in his cohort. Feng et al 34 also observed these findings and developed a more graded and sensitive assessment of corticospinal tract lesion load (ie, weighted corticospinal tract lesion load) that could effectively predict the motor outcomes at 3 months after stroke. If the lesion load cumulates over 7.00cc, it reaches a threshold signifying that corticospinal tract damaged to a critical unrecoverable point. None of the patients in the two independent cohorts whose lesion load greater than 7.00 cc recovered meaningful motor function. Again, the dosage of therapy did not have a significant correlation with degree of motor recovery. Interestingly enough, proportional recovery rule has also been observed in poststroke language recovery37 as well as visuospatial neglect recovery.38
Early mobilisation after stroke
The critical period to commence of rehabilitation after stroke has been poorly defined due to the complexity of trials at an early phase of stroke. Although early mobilisation has been recommended in many guidelines, limited evidence is available to guide the practice of rehabilitation. The international, multicentre A Very Early Rehabilitation Trial(AVERT) phase III study 39 reported by Bernhardt et al randomised 2104 patients from 56 sites in 5 countries to compare a frequent, higher dose of out-of-bed mobility based training within 24 hours stroke to usual care alone. Surprisingly, a higher dose protocol resulted in lower odds of a favourable outcome at 3 months, which contradicted the traditional belief that ‘more is better’. Interestingly, subsequent40 prespecified analyses suggested higher frequency of intervention is associated with more favourable outcome. The results provided a new research direction regarding the training frequency and intensity at the very early period of stroke, which has been simply-out-of-bed protocol. The feasibility and safety of 30 min twice daily, 5-day treadmill training, starting within 48 hours symptom onset was investigated in 20 patients who had a acute stroke by Strømmen et al.41 An intensity of >50% of the individual heart rate reserve was obtained in 31% of training sessions. This study showed that early intensive treadmill training in patients who had a acute stroke is possible, and thus did not lend support to the concerns surrounding specific training programmes in the very acute phase of stroke.
The promise of stem cell therapy
If the motor tract is entirely or critically damaged or there is a lack of neurosubstrate to recover, inducing exogenous stem cells or bypassing the damaged the motor tract with brain–computer interface technology appears to be a logical next step. Despite all the exciting basic science discoveries made in stem-cell research highlighted above, there is still a lack of a breakthrough in this area in human study. Steinberg et al 20surgically transplanted modified bone marrow-derived mesenchymal cells to 18 patients who had a stroke to test the safety and preliminary efficacy. In an open label single-arm two-centre study, 16 (out of 18) patients completed 12 months follow-up visit. All 18 patients experienced one or more treatment-emergent adverse events including six serious ones although none of them left a sequel. The average improvement on the National Institutes of Health Stroke Scale (NIHSS) is 2 points—an improvement smaller than the minimal clinically important different (MCID) of 4 points, but the improvement on Fugl-Meyer Motor Function (FMMF) Scale is 11.4 points, and is above the MCID which ranges from 4.3 to 7.3 points in patients who had a chronic stroke.42
Brain and peripheral stimulation
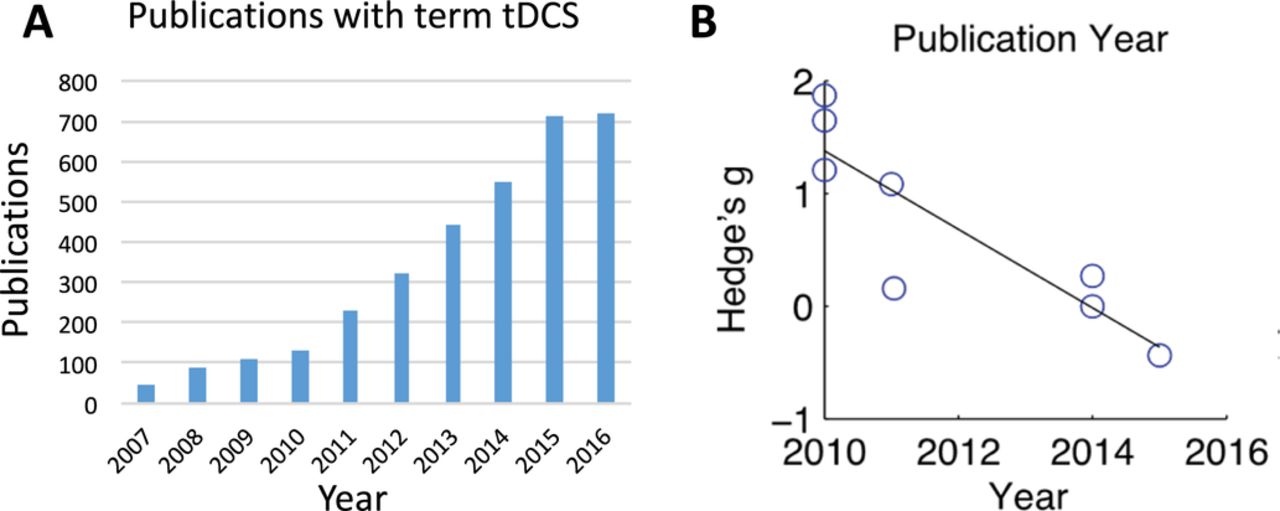
Transcranial direct current stimulation(tDCS), a non-invasive brain stimulation tool, remained a hot topic in stroke recovery and it has been investigated for motor, language, dysphagia, visual field deficit, etc.43 In prior studies, stimulating specific domain holds promise in reducing domain-specific deficit; however, stimulating an ancillary system (the primary motor system) that is functionally connected to the residual language network also improved naming significantly in patients who had a stroke in a recent study.44 In this study, 26 patients with chronic aphasia were randomised to a sham group and a stimulation group with anodal stimulation being administered to the left primary motor cortex twice daily at the beginning of each speech training session. Each group received a highly intensive naming therapy twice daily with each session lasting 90 min over a 2-week period (total 8 days). Despite the plethora of manuscripts published in the last decade (figure 1), results continued to be mixed and inconsistent. One possible explanation is a lack of establishment of a dose–response relationship (or defining the optimal dose). Chhatbar et al 45 revealed a positive dose–response relationship between the current density (current/pad size) and the reduction of motor impairment (measured by Fugl-Meyer Upper Extremity Scale) by a meta-regression of eight published studies with various current level and pad size. If a higher current proves to be effective and safe, it may have a profound impact on the tDCS stroke recovery field. Reverse translational research is urgently needed as tDCS animal studies are scarce and several dosing-related questions can only be addressed in animals.

{kind=link}
(A) Shows the number of manuscripts from 2007 to 2016 if searching Pubmed with term of ‘transcranial direct current stimulation (tDCS)’; (B) shows the changes of effect size (ie, Hedge’s g) of tDCS from poststroke upper extremity motor recovery trials over the years. The values of Hedge’s g are from the able 2 in Dr Chhatbar’s study.45
The swallowing treatment using pharyngeal electric stimulation (PES) trial was an international, multicentre, randomised, sham-controlled, patient-masked, outcome assessor-masked, parallel-group trial.46 In this trial, 162 subacute ischaemic or patients who had a haemorrhagic stroke with dysphagia (defined as a penetration aspiration score of ≥3 on videofluoroscopy) were randomised to receive three consecutive days of PES or sham treatment. Although PES was shown to be safe without any serious device-related adverse events, the primary outcome (radiological aspiration at 2 weeks assessed by penetration aspiration score) was not separated between the two groups. The authors believed that underdosing of PES is one of the potential factors contributing to the neutral result.
Summary
Stroke recovery and rehabilitation is a dynamic scientific field. 2016 is an exciting year for scientists and patients, with several breakthroughs in both benchwork research as well as the clinical research, as highlighted above. Looking into 2017, the priorities of stroke recovery should be: (1) to better understand the pathophysiology and factors underlying the recovery trajectory,47 especially in the first 90 days after stroke during which the majority of improvement occurs; (2) to identify the novel targets; (3) to maximally preserve, protect and recover brain function by harnessing brain plasticity; (4) to ensure basic and translational science research are closely engaged and to address the unmet needs in clinical practice and (5) to translate restorative poststroke therapeutics through collaborative, rigorous, transparent research efforts.
Acknowledgments
We thank Dr Pratik Chhatbar for his assistance on generating the figure 1, and Drs James Sawers and Alexandra Parashos for their generous comments and edits on the manuscript. Dr WF acknowledges his grant supports from National Institute of Health (P20GM109040) and American Heart Association (14SDG1829003).
References
Footnotes
Contributors HZ and WF contributed to the concept and design of the study. HZ, NC and YY drafted and edited different sections of the manuscript. WF oversaw the manuscript, made critical edits and finally approved the manuscript. All authors read and approved this submitted manuscript.
Competing interests None declared.
Provenance and peer review Commissioned; externally peer reviewed.
Correction notice This article has been corrected since it published Online First. The heading ’Early immobilisation after stroke' has been corrected to ’Early mobilisation after stroke'.