Article Text

Abstract
Background The brain-gut axis is a major regulator of the central nervous system. We investigated the effects of treatment with broad-spectrum antibiotics on gut and brain inflammation, infarct size and long-term behavioral outcome after cerebral ischemia in rats.
Methods Rats were treated with broad-spectrum antibiotics (ampicillin, vancomycin, ciprofloxacin, meropenem and metronidazole) for 4 weeks before the endothelin-1 induced ischemia. Treatment continued for 2 weeks until the end of behavioral testing, which included tapered ledged beam-walking, adhesive label test and cylinder test. Gut microbiome, short-chain fatty acids and cytokine levels were measured together with an assessment of infarct size, neuroinflammation and neurogenesis.
Results The results revealed that the antibiotics exerted a clear impact on the gut microbiota. This was associated with a decrease in systemic and brain cytokine levels, infarct size and apoptosis in the perilesional cortex and improved behavioral outcome.
Conclusion Our results highlighted the significant relationship between intestinal microbiota and beneficial neuro-recovery after ischemic stroke.
- Stroke
- Stroke Rehabilitation
- Cerebral Infarction
Data availability statement
Data are available upon reasonable request. The data contains the original images as well as the original files, the raw data are available from the authors CL and CZ (leo_liuchang@163.com and cszhao@cmu.edu.cn). Permission to reuse the data need to be granted with the permission of the journal and the corresponding author.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key messages
What is already known on this topic
The modulation of the gut microbiota and brain resident immune cells could act as a major factor in the process of pathophysiology of stroke. How to modify the structure of the microbiota in intestine to ameliorate the outcome of ischaemic stroke has received extensive attention.
What this study adds
In our study, a combinatorial antibiotic is applied to modify the structure of the flora after stroke, thereby regulating the inflammatory reaction through the peripheral circulation and central nervous system and ultimately ameliorate the outcome of stroke.
How this study might affect research, practice or policy
This article may provide new avenues for further exploring mechanisms of brain–gut axis as well as promising therapeutic strategies for antibiotic application and to improve stroke recovery.
Introduction
Faecal microbiota transplantation, treatments with broad-spectrum antibiotics and the provision of probiotics are some of the approaches which have been exploited to examine the relationship between gut microbiota and brain diseases. A healthy gut microbiota is necessary for human welfare, including the functions of the immune system and central nervous system (CNS).1 The gut microbiota not only protects the host against invading pathogens, but also stimulates angiogenesis, regulates fat storage and maintains intestinal permeability.2 Gut microbiota disorders are associated with a variety of diseases, for example, inflammatory bowel disease, obesity and type 1 diabetes, and nervous system diseases such as stroke, multiple sclerosis, Parkinson’s disease, Alzheimer’s disease, anxiety and neurodevelopmental disorders.
The relationship between the intestinal flora and stroke has attracted considerable attention. The destruction of the epithelial barrier after a stroke makes the intestine a potential gathering site for invasive pathological bacteria. In addition, a stroke changes the activity of the autonomic nervous system, which not only contributes to immune suppression but also affects bowel motility and permeability.1 The products released from the intestinal tract and the flora can gain access to the circulation and cross the leaky blood–brain barrier (BBB) to disturb the regulation of the immune response in the CNS.3 Thus, it is not surprising that it has been speculated that antibiotics may affect the outcome of stroke. Indeed, administration of different antibiotics (ie, ampicillin, metronidazole, neomycin sulfate and vancomycin) for 4 weeks before an experimental stroke greatly reduced the infarct volume and restored the sensorimotor function of mice, this effect possibly being mediated through the modification of the gut flora.4 However, extended administration of antibiotics may also lead to sepsis and severe bloody diarrhoea caused by reduced gut motility and increased permeability and eventually impaired neurological function.5 Nonetheless, exactly how modulation of the gut microbiota affects the evolution, development and recovery after a stroke is not completely understood.
Therefore, we aimed to explore whether an intestinal microbial shift caused by broad-spectrum antibiotics could alter brain pathology and stroke recovery. We hypothesised that the changes in the bacterial populations induced by antibiotics would trigger a peripheral immune reaction, which in turn would result in neuronal protection and functional restoration after a stroke. In these experiments, rats were divided into sham-operated (SHAM) group, ischaemic (ISC) group and ischaemic rats treated with a combination of antibiotics (ISCAB) group. We found that the combined broad-spectrum antibiotics selectively affected the abundance of the intestinal flora after stroke, which could lead to an increase in the intestinal content of short-chain fatty acids. In addition, antibiotics reduced the inflammatory response in the intestine and modulated the function of microglia resulting in decreased neuronal apoptosis and a reduced infarct volume consequently leading to an improved behavioural recovery.
Materials and methods
Animals and experimental procedures
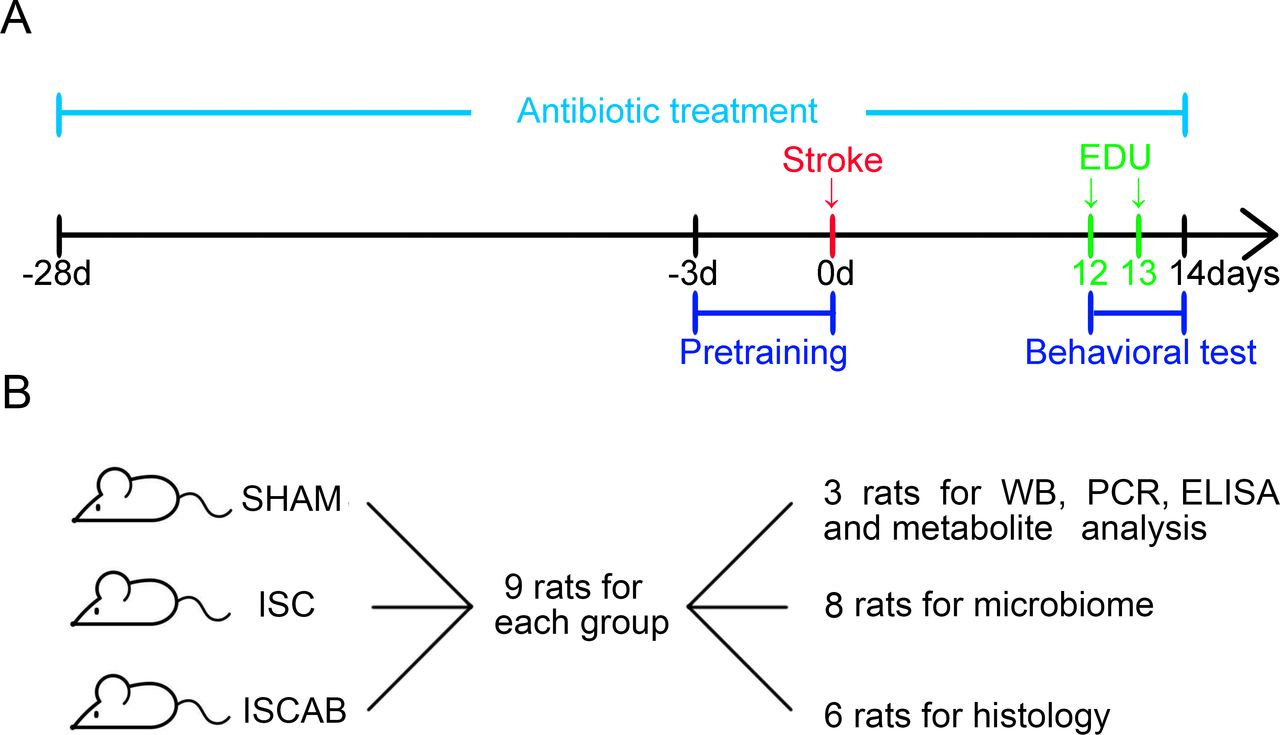
The experimental design is shown in figure 1. Male Wistar rats (220–280 g, age: 4 weeks) were obtained from China Medical University (Shenyang, China). Rats were housed in a light-controlled, temperature-controlled and humidity-controlled environment with free access to food and water. The rats were randomly divided into three groups based on a random number table generated by Microsoft Office Excel 2019: sham-operated rats (n=9, SHAM), rats subjected to cerebral ischaemia (n=9, ISC) and rats with ischaemia treated with broad-spectrum antibiotics for 4 weeks before the ischaemic operation and continued until the rats were sacrificed (n=9, ISCAB). The quintuple cocktail of antibiotics consisted of ampicillin (1 g/L), vancomycin (500 mg/L), ciprofloxacin (200 mg/L), meropenem (250 mg/L) and metronidazole (1 g/L) delivered in the drinking water.6 Body weight was monitored every day during the experiment (online supplemental figure 1). The mortality rate was less than 15% during surgery. There was no loss due to post-operation complications such as dysentery nor signs of sepsis caused by long-term antibiotic use. The ISCAB rats cannot be blinded to the investigator due to their watery faeces. A detailed description of the materials and methods used is provided in the online supplemental material.
Supplemental material

Experimental design. (A)The arrows show the timing of pretraining, stroke, EDU injection and sacrifice. Antibiotics were given in the drinking water for 4 weeks before ischaemia and the treatment was continued until the end of the experiment. (B)Number of rats used for behaviour, histology, WB, RT-qPCR and ELISA and microbiome and metabolites analysis. EDU, 5-ethynyl-2'-deoxyuridine; ISC, ischaemic; ISCAB, ischaemic rats treated with a combination of antibiotics; SHAM, sham-operated; WB, western bolt.
Results
The administration of antibiotics causes a shift in the gut microbiota after a stroke
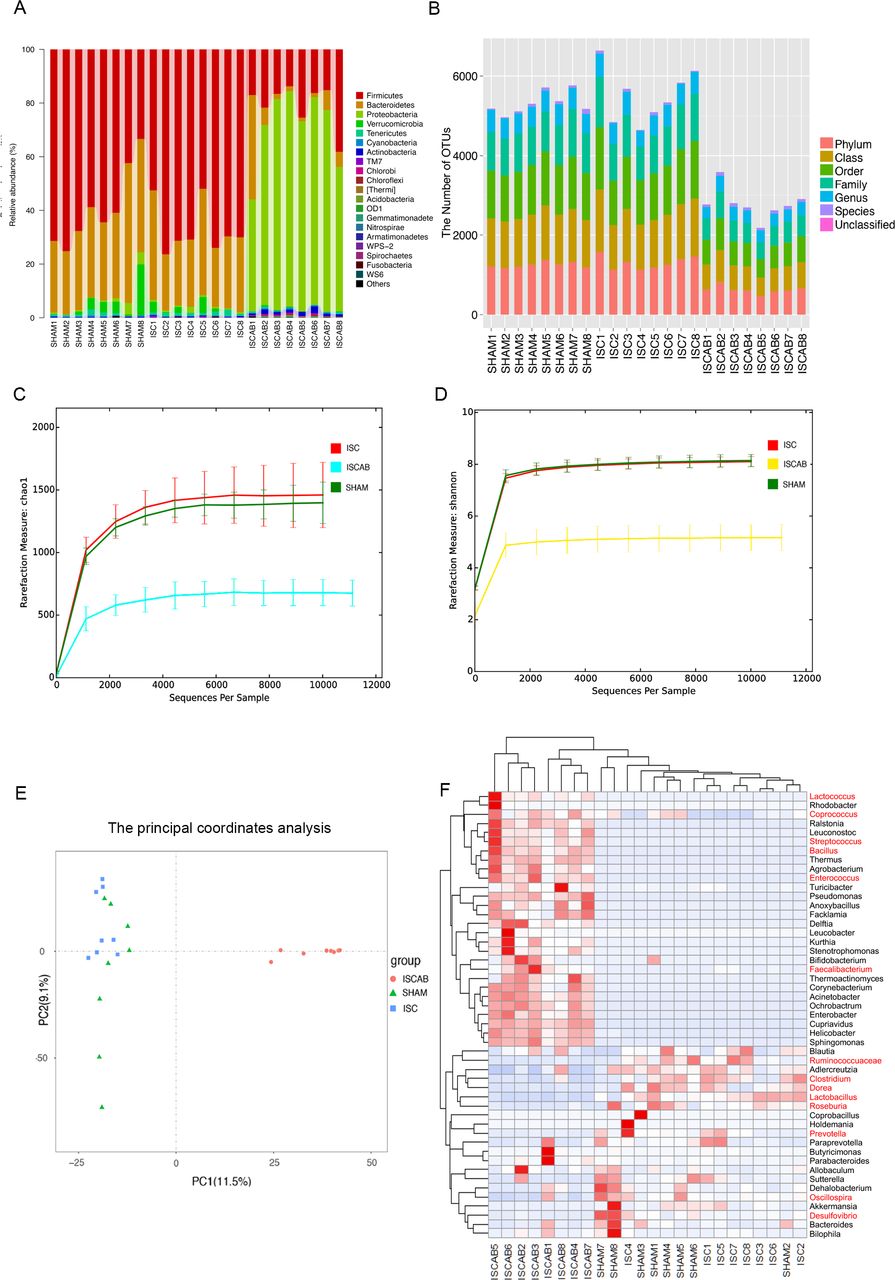
We performed 16S rRNA sequencing of the intestinal contents collected from the experimental rats after sacrifice (n=24). The relative abundance of gut microbiota at the phylum level showed that Firmicutes had the highest relative abundance in the ISC rats (67.1%±3.3%) and antibiotics expanded the members of Proteobacteria, which occupied the dominant position of the gut microbiota in the ISCAB group (68.1%±4.8%) (figure 2A). The practice of clustering operational taxonomic units (OTUs) at 97% 16S sequence identity is intended to group together functionally diverse lineages. The results suggested that the abundance of microbiota colonies in the gut had been reduced by antibiotics (figure 2B). Subsequently, the Chao1 Index and Shannon Index were analysed based on the number of OTUs. These results showed that the microbial α-diversity in the ISCAB group was lower than other groups, indicating that the intervention with the antibiotics had indeed decreased the microbial α-diversity and richness (figure 2C and D). The principal coordinates analysis (PCoA) scatterplot was performed to examine if there was any difference between the three groups based on the relative abundance of OTUs in terms of the collected ileal contents. The ISCAB group was clearly distant from the other groups indicating that the flora composition of ISCAB rats was different from the other two groups (Principal component 1 (PC1) explained 11.5% and Principal component 2 (PC2) explained 29.1% of the trend) (figure 2E).

Overall taxonomic analysis, microbial community diversity analysis and heatmap analysis. (A)Taxonomic composition and distribution map of flora were summarised at the phyla level. (B)OTUs in each sample that can be classified to the level of the phylum, the class, the order, the family, the genus and the species. (C)Measure of α-diversity species of samples by Chao1 Index based on the number of rare species. (D)α-diversity of samples indicated by Shannon’s diversity analysis. (E)β-diversity of principal component analysis. PCoA of gut microbiota indicated the microbial composition. (F)Hierarchically clustered heatmap analysis of the top 50 bacteria at the genus level in gut (n=8 per group). ISC, ischaemic; ISCAB, ischaemic rats treated with a combination of antibiotics; PCoA, principle coordinate analysis; SHAM, sham-operated.
Although the diversity in the ISCAB group had declined due to antibiotic treatment, the results obtained from hierarchical clustering and heat map analysis showed that absolute amount of probiotics such as Lactococcus, Coprococcus, Bacillus, Streptococcus, Enterococcus and Faecalibacterium had increased in the ISCAB rats (figure 2F and online supplemental figure 2A). The proportion of Desulfovibrio, which is a pathogenic bacterium, decreased in ISCAB rats and antibiotics also reduced the proportions of Ruminococcaceae, Clostridium, Dorea, Lactobacillus, Roseburia, Oscillospira and Prevotella (online supplemental figure 2A). These reduced microbiota are commonly considered to produce butyrate in the ileum.7–13
Antibiotics alter the concentrations of SCFAs
Given that short-chain fatty acids (SCFAs) produced by bacterial fermentation are important metabolites in the intestine and many of them have immunomodulatory functions, we quantified the intestinal metabolites after the antibiotic intervention. Compared with ISC rats, the concentration of acetate increased (105.6±10.1 µg/g vs 58.9±5.0 µg/g, p<0.05), whereas that of butyrate decreased (0.1±0.03 µg/g vs 0.27±0.04 µg/g, p<0.05) after the antibiotics (online supplemental figure 2B).
Since acetate14 and butyrate15 are specifically produced by the microbiome, we correlated the changes in the microbiota with that of their metabolites. The results indicated that the levels of acetate were significantly correlated with Blautia and Turicibacter in SHAM rats, with Adlercreutzia, Bacteroides, Butyricimonas, Paraprevotella, Clostridium, Sutterella, Dehalobacterium, Oscillospira and Parabacteroides in ISCAB and with Ruminococcus both in ISC and ISCAB rats. The levels of propionate were significantly correlated with Christensenella and Ruminococcus in SHAM rats. The levels of butyrate were significantly correlated with Ruminococcus in ISC rats. The levels of caproate were significantly correlated with Cupriavidus in ISC (p<0.05) (online supplemental figure 3). Online supplemental figure 4 shows the Pearson correlation between the top 30 microbiota and behavioural tests. It is worth noting that in this analysis, Bacteroides, Butyricimonas, Christensenella, Dehalobacterium, Oscillospira, Parabacteroides, Paraprevotella and Ruminococcus showed the negative correlations with behavioural indices of rat forelimbs. This is consistent with the previous results of acetic acid, suggesting that the changes of acetic acid produced by the gut microbiota might play an important role in the behavioural recovery. The combined data clearly revealed that a major effect of antibiotic treatment had been to alter the diversity and abundance of gut microbiota and their secretion of SCFAs.
Decreases in systemic inflammatory response by antibiotics
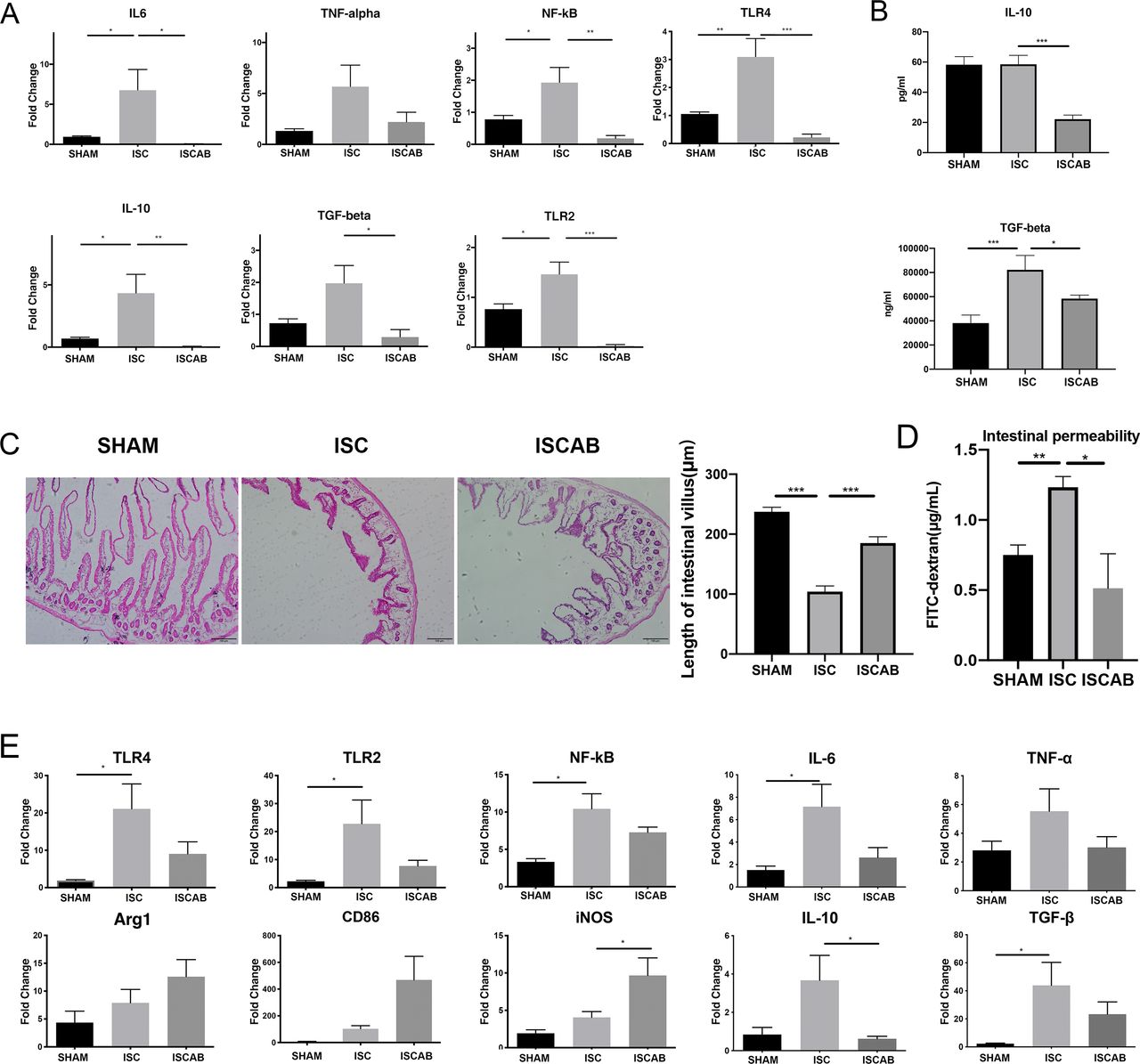
SCFAs have been recognised as negative regulators to the gut inflammation after cerebral ischaemia. Thus, the expression of proinflammatory (Toll-Like Receptor 4 (TLR4), Toll-Like Receptor 2 (TLR2), Nuclear factor kappa B (NF-kB), interleukin 6 (IL-6) and tumour necrosis factor α (TNF-α)) and anti-inflammatory cytokines (IL-10 and Transforming growth factor beta (TGF-β)) mRNA was investigated in intestinal tissue. We found that in ISC rats, levels of all the above cytokines except TNF-α and TGF-β increased as compared with SHAM rats and the microbiota changed by antibiotics had opposite effects on the amounts of both anti-inflammatory and proinflammatory markers. However, although TNF-α showed the same tendency, there is no significant difference between the three groups (figure 3A). When TGF-β and IL-10 levels were measured in blood, we found that both markers had decreased after antibiotic treatment and TGF-β showed significant increase in ISC group compared with SHAM group (figure 3B).
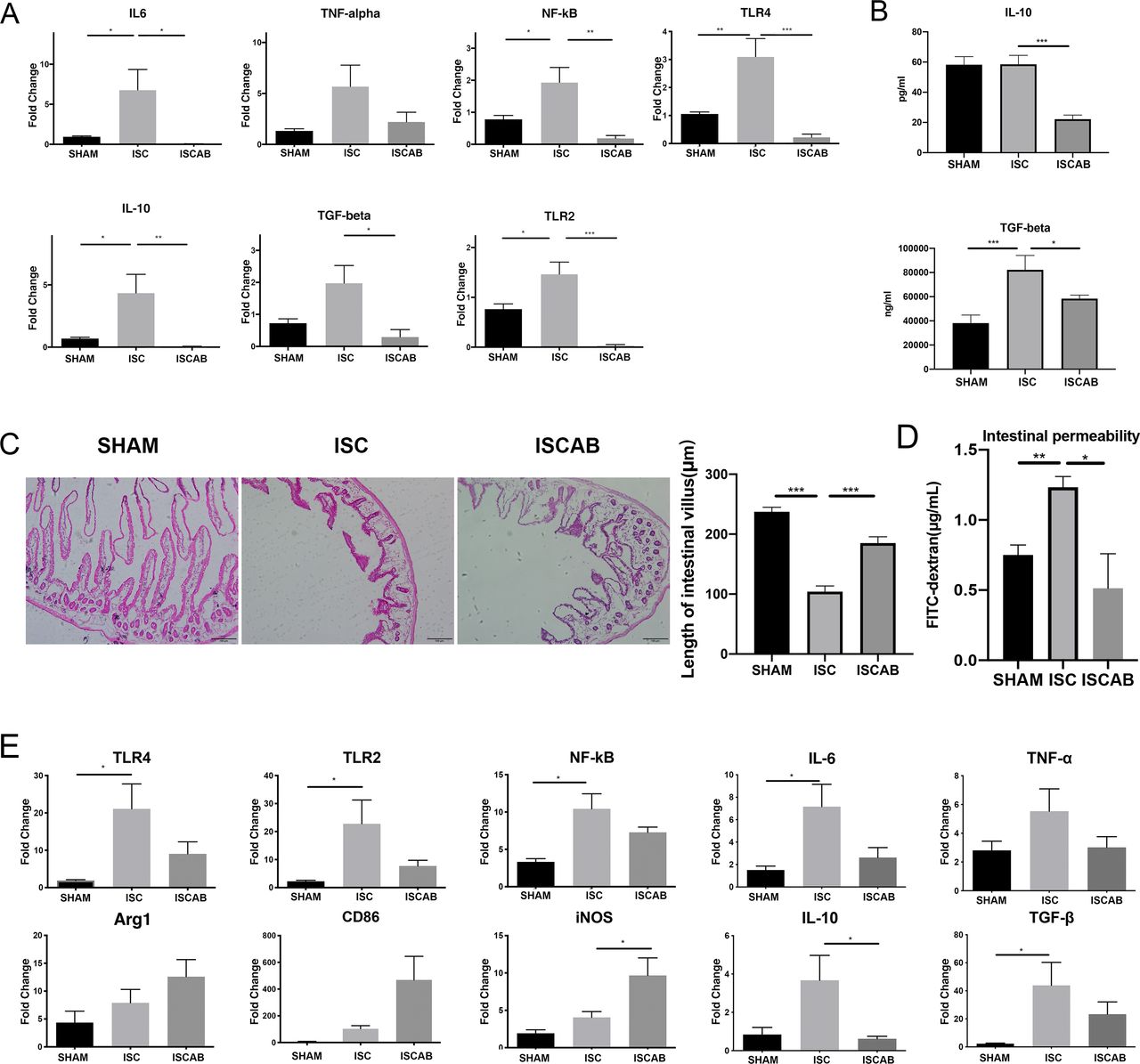
Effect of cerebral ischaemia and antibiotic treatment on cytokine levels. (A)Relative mRNA expressions of NF-kB, TLR2, TLR4, IL-6, TNF-α, IL-10 and TGF-β in the gut of the rats. (B)Concentrations of plasma TNF-β and IL-10. (C)Average length of intestinal villus. (D)Serum levels of fluorescein-labelled isothiocyanate-dextran (FITC-dextran) indicating intestinal permeability differences. (E)Relative mRNA expression of CD86, Inducible nitric oxide synthase (iNOS), Arginase 1 (Arg1) and inflammatory-related factors NF-kB, TLR2, TLR4, IL-6, TNF-α, IL-10 and TGF-β in the perilesional cortex. Data are mean±SEM, n=3 per group. Statistical significance: *p<0.05; **p<0.01; ***p<0.001. IL-6, interleukin 6; IL-10, interleukin 10; ISC, ischaemic; ISCAB, ischaemic rats treated with a combination of antibiotics; SHAM, sham-operated; TNF-α, tumour necrosis factor α.
The disruption of intestinal mucosa was specifically reflected in the decreased length of small intestinal villi, the reduced crypt foci and the decreased thickness of intestinal wall. In our experiments, we observed significant changes in the morphology of the small intestinal villi. The mean length of small intestinal villi was significantly reduced after the ischaemic event (p<0.001), and when treated with a long course of antibiotics, the length of small intestinal villi was significantly restored compared with that in the ISC group (p<0.001, figure 3C), and we found that the ISCAB group showed a significant increase in intestinal permeability to FITC-dextran when compared with the ISC group (p<0.05, figure 3D), suggesting the recovery of mechanical barrier of intestinal mucosa after treatment.
Then we used Quantitative reverse transcription PCR (RT-qPCR) to examine if there were alterations of microglia-associated markers and cytokines in the perilesional cortex. There is a trend for the expression of Arginase 1 (Arg1), CD86 and Inducible nitric oxide synthase (iNOS) to increase in ISC rats and a further increase in the ISCAB group; however, only the increase of iNOS in the ISCAB group reached statistical significance (p<0.05, figure 3D). In ISC group, the mRNA levels of TLR4, TLR2, NF-kB, IL-6 and TGF-β showed significant increases as compared with the SHAM group. These increases tended to be reversed in the ISCAB group, although the change did not reach statistical significance. IL-10 and TNF-α also showed the tendency to increase in ISC but the change did not reach statistical significance. Nonetheless, in the ISCAB group, IL-10 mRNA level was significantly lower than that in the ISC group (p<0.05, figure 3E).
Improved behavioural outcome following stroke by antibiotics
Next, we aimed to evaluate how the antibiotic treatment affected neurological deficit in chronic post-stroke phase. Sensorimotor functions were measured on postoperative day 14 to assess behavioural outcomes in ischaemic rats. In the beam walking test, both forelimb and hindlimb slips were increased by ischaemia (8.7%±2.0% vs 26.3±3.7%, p<0.001; 21.8±4.2% vs 49.6±2.8%, p<0.001, figure 4A) and the impaired hindlimb function was reversed in ISCAB group when compared with ISC group (17.0%±2.3% vs 26.3±3.7%, p<0.05; 26.9±5.1% vs 49.6±2.8%, p<0.01, figure 4A). In the sticky label test, both the time taken to first touch the adhesive tape (14.3±1.6 s vs 19.2±1.5 s, p<0.05) and its removal time (44.4±5.1 s vs 75.1±9.6 s, p<0.05, figure 4A) were reduced in the ISCAB group as compared with the ISC group. Ischaemia impaired spontaneous forelimb use in cylinder test, and this was reversed by antibiotics (40.7%±2.9% vs 20.1±4.7%, p<0.01, figure 4A). All the behavioural results together revealed that the sensorimotor functions of rats in the ISCAB group had undergone a significantly improved recovery after a stroke.

Effect of cerebral ischaemia and antibiotics on behavioural performance, infarct ratio and apoptosis in the perilesional cortex. (A)Improved behavioural outcome in the group receiving antibiotics, n=9 per group. (B)Cresyl violet staining reveals a typical infarct and neuroprotection by antibiotics, n=6 per group. (C)Western blotting images and data of Bax and Bcl2, n=3 per group. (D)Representative images and data for TUNEL/+NeuN cells in the perilesional cortex, n=6 per group. Data are mean±SEM. Scale bar=50 µm. Statistical significance: *p<0.05; **p<0.01; ***p<0.001. ISC, ischaemic; ISCAB, ischaemic rats treated with a combination of antibiotics; SHAM, sham-operated; GAPDH, Glyceraldehyde 3-phosphate dehydrogenase.
Antibiotics cause a decrease in infarct volumes and perilesional apoptosis
The endothelin-1 induced experimental stroke caused extensive damage to the cortex and striatum. To examine whether antibiotics decreased the extent of the ischaemic injury, infarct volumes were analysed from cresyl violet-stained sections. ISCAB rats had significantly smaller infarct volumes as compared with ISC rats (6.2%±0.8% vs 11.8±1.4%, p<0.001, figure 4B).
Bax, as a marker of apoptosis, was significantly decreased in the ISCAB rats compared with their ISC counterparts (figure 4C). However, there was no difference in the expression of Bcl2 in the different experimental groups (figure 4C). NeuN can be used as a neuronal marker, which we used to identify the type of cells undergoing apoptosis. We found that there was a significant overall group effect in the number of the Tunel+/NeuN+cells in the perilesional cortex (p<0.001). The number of Tunel+/NeuN+cells in the ISC group was increased compared with that in the SHAM group (p<0.001, figure 4D). After 4-week administration of antibiotics, the number of Tunel+/NeuN+cells was decreased compared with that in the ISC group (p<0.01, figure 4D). These results confirm that antibiotic can reduce apoptosis of neuronal cells.
Changes in glial and microglial cells after cerebral ischaemia
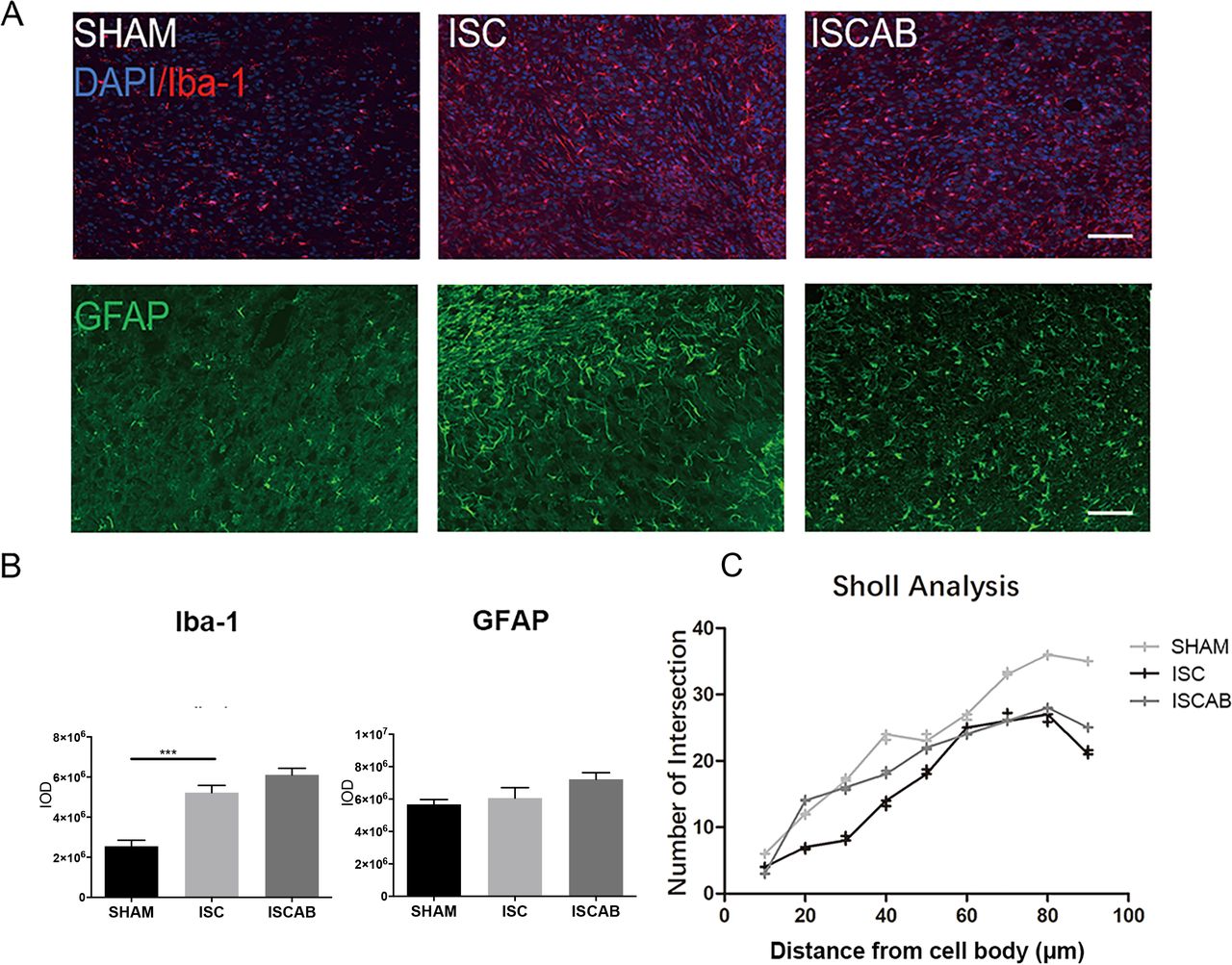
There was a significant overall group difference in the number of Iba-1+cells in the perilesional cortex (figure 5A,B). In ISC animals, the expressions of Ionized calcium binding adaptor molecule 1 (Iba-1) and Glial fibrillary acidic protein (GFAP) were higher than in the SHAM rats (p<0.05); however, this effect was not reversed by antibiotics. Sholl analysis showed a reduction in the total branch number and length of microglia in the perilesional cortex after ischaemia (figure 5B). However, the ISCAB rats did not differ from their ISC counterparts (figure 5C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Gut inflammation response in the perilesional cortex. (A)Fluorescence images and quantification of Iba-1+cells and GFAP+cells in the perilesional cortex, n=6 per group. (B)Western blotting and quantification of Iba-1 and GFAP in the perilesional cortex, n=3 per group. (C)Quantification of the number of microglia branching, n=3 per group. Data are mean±SEM. Scale bar=50 µm. Statistical significance: *p<0.05; ***p<0.001. ISC, ischaemic; ISCAB, ischaemic rats treated with a combination of antibiotics; SHAM, sham-operated; DAPI, 4,6-diamidino-2-phenylindole dihydrochloride.
Discussion
Changes in the intestinal flora are reflected in SCFA metabolism
Previously, it has been demonstrated that rats develop an abnormal microbial composition after stroke and this in turn affects the prognosis of stroke. Oral administration of antibiotics have been reported to decrease the volume of cerebral infarct and reduce cerebral oedema by changing the intestinal flora.4 16 The absence of microbiota within 1–4 days after stroke did not affect infarct size and increased mortality within 5–7 days in mice, and this could be prevented by continuous antibiotic therapy or transplantation of gut microbiota obtained from Specific-pathogen-free (SPF) animals.5 In our study, the proportion of beneficial bacteria such as Lactococcus, Coprococcus, Bacillus, Streptococcus and Faecalibacterium was significantly upregulated after treatment with antibiotics. After depletion of a large number of antibiotic-sensitive flora by the antibiotic cocktail, the other flora are no longer subjected to their competitive inhibitory effect, which can represent a growth advantage.17 Part of the intestinal flora was resistant to the antibiotic treatment. For example, Enterococcus is known to be resistant to ampicillin to a certain extent.18 Antibiotic resistance of Bacillus is another example.19 There is previous evidence that a probiotic intervention, including Lactococcus and Coprococcus, significantly ameliorated depression-like symptoms and improved cognitive functions of rats.20 The increased relative abundance of Streptococcus and Faecalibacterium is known to attenuate the symptoms of colitis and suppress the immune response.21
Short-chain fatty acids are important metabolites of intestinal flora. Acetate and butyrate are the most abundant SCFAs in the intestinal tract (≥95%).22 Acetate can serve as a carbon source for gut microbiota, and help to maintain the integrity of the epithelial barrier by regulating the properties of tight junction proteins. Acetate can be produced by most of the enteric bacteria such as Lactobacillus spp, Prevotella spp, Ruminococcus spp, Streptococcus spp and Blautia spp,14 23 whereas butyrate is produced by Clostridium leptum, Faecalibacterium, Oscillospira and Coprococcus.15 24 Overall, our results revealed an increase in the proportions of the above microflora in conjunction with the changes of intestinal SCFAs.
SCFAs participate in the inflammatory regulation that affects the CNS via the circulation
Brain ischaemia leads to the breakdown of mucosal barriers and bacterial translocation. Microbial components, such as bacterial cell components or toxins, can enter the host’s blood circulation or lymphatic organs, further increasing the intestinal permeability and reducing the immune response in peripheral circulation, ultimately leading to bacterial translocation and infection.25 In experimental stroke, T and B cell numbers are significantly reduced in Peyer’s patches.26 Immune cells also can infiltrate across the leaky BBB into the parenchyma at the site of the ischaemic lesion.
SCFAs can repair the intestinal barrier. For example, acetate can increase cell membrane assembly, mucosal cell migration and the proliferation and differentiation of colonocytes as well as modulating the various biological responses of the host, for example. reducing inflammation and oxidative stress.27 In vitro, low concentrations (1–10 mM) of sodium butyrate significantly could improve the transepithelial resistance and permeability of the epithelium of colonic cells.28 Similarly, increases in the circulating levels of SCFAs may induce therapeutic effects during the chronic post-stroke recovery period. SCFAs are especially important for the human body in its defence against intestinal diseases such as Crohn’s disease, ulcerative colitis and colorectal cancer.15 In general, two different mechanisms are involved in these processes; the first is associated with a direct activation of certain G-protein-coupled receptors and the second is attributable to the direct inhibition of nuclear class I histone deacetylases (HDACs).15 The inhibition of HDACs is mainly associated with the anti-inflammatory immune phenotype, including declines in the concentrations of inflammation-associated cytokines (IL-6, IL-8 and TNF-α), reducing NF-kB activity and avoiding pathogen expansion.29 TNF-α expression was found to be downregulated after a diet containing 5% Plantago ovata seeds seeds compared with control animals, while TNF-α and NF-κB expression in the intestine was downregulated after feeding with a diet containing 8% oligofructose-enriched inulin. In both studies, it was confirmed by two studies that the reduction of intestinal inflammatory factors was associated with higher concentrations of butyrate.30 31 This can be attributed to the anti-inflammatory effects of butyrate mediated by the inhibition of NF-κB signalling pathway and HDAC. As butyrate has been reported to bind and activate the nuclear transcription factor Peroxisome proliferator-activated receptor gamma (PPARγ), thereby antagonising NF-κβ signalling.32 33 Our results revealed that the amounts of NF-kB, TLR2, TLR4, IL-6, TNF-a, IL-10 and TGF-β decreased in gut, which means that the extent of intestinal inflammation had been indeed significantly reduced by antibiotics. However, IL-10, as a recognised anti-inflammatory factor, is reduced significantly. This may result from the fact that SCFAs can inhibit Lipopolysaccharide-induced IL-10 production as previous shown.34 Taken together, it seems that SCFAs exert a variety of beneficial functions in ischaemic stroke.
Antibiotics promote a recovery of neurological function
Our results show that the gut microbiota has an important impact on the behavioural recovery taking place after an experimental stroke. In addition, administration of these antibiotics affected the gut microbiome leading to neuroprotection and an improved functional recovery in ISC animals. Benakis et al showed that infarct volumes were reduced by 60%±6% in antibiotic-treated mice with an altered intestinal flora in comparison to antibiotic-resistant mice and this was associated with better sensorimotor functions.35 There is also evidence that apoptotic cell death may be associated with impaired survival of newly formed neurons.36 In line with these findings, our study showed that the animals drinking the antibiotic cocktail showed less evidence of apoptotic cell death in the perilesional brain. Although several mechanisms are likely involved in the bidirectional communications of the microbiota–gut–brain axis following a stroke, one important consideration is the initiation of the brain inflammatory response by the gut microbiota in conjunction with the cooperative role of microglia.35
Microglia activation by stroke and antibiotics
Proinflammatory mediators released from microglia have been associated with the neuroinflammatory injury evident after a stroke. On the other hand, microglia contribute to neurogenesis and guidance of sprouting vessels through the production of neurotrophic factors, phagocytosize synapses and reshaping neuronal circuitries. Interestingly, microglia, unlike peripheral macrophages, may be more susceptible to changes in the microbiome.37 Erny et al found that in comparison to SPF mice, germ-free (GF) microglia expressed reduced mRNA levels for several activation markers (eg, Il-1α, Stat1, Jak3 and B2m). In addition, the expressions of other genes controlling cell proliferation, cell cycle and apoptosis (eg, Cdk9, Ccnd3 and Bcl2) were substantially increased in GF mice. Accordingly, an immature morphology and also an increased microglial density were observed in different brain regions of GF mice. GF mice exhibited an upregulation of the surface markers CSF-1R, F4/80 and CD31, proteins which are known to be downregulated in mature adult microglia. Furthermore, GF microglia displayed a reduced capacity to respond to a viral infection challenge. A reduced complexity of microbiota also leads to microglial defects after antibiotic use. In contrast, the re-colonisation of the complex flora and SCFAs treatment restored the microglial characteristics.37
Our immunofluorescence and RT-qPCR results in the cerebral cortex detected signs of upregulation of microglia-related genes after stroke. We noted that the usage of antibiotics did not significantly affect the number of Iba-1+cells but increased the number of proinflammation microglia, which means the ratio of the microglial phenotypes indeed changed. Sholl analysis showed that the alterations in microglial morphology induced by oral antibiotic therapy are indicative of the dynamic polarisation states of these cells. Activated microglia can retract their processes and enlarge their cell bodies in a manner like activated phagocytic cells, increasing the relative areas of Iba-1 staining after a stroke, while ameboid microglial demonstrate a protective state, attenuating the damage associated with the neuroinflammation associated with a stroke. The anti-inflammatory gene expression profile of microglia was associated with reduced mRNA expressions of TNF-α and IL-1β but increased expressions of TGF-β1 and IL-10.38 In our study, the levels of an activated microglia-related marker increased in parallel to the findings of reduced apoptosis. Although there was an increase in proinflammatory microglia, as indicated by the upregulation of the proinflammatory enzyme iNOS, there was also evidence of downregulation of both inflammatory and anti-inflammatory cytokines in the cortex at the same time, which hinted that the inhibition of the immune response may due to dysfunction and immaturity of microglial cells caused by the broad-spectrum antibiotic.37 This change in the properties of the microglia could lead to a reduction in the inflammatory response in the brain after stroke and promote stroke recovery.
Limitations
However, our study has some limitations. First, antibiotics may cross the leaky BBB. Metronidazole and meropenem are more likely to enter the CNS and affect the function of microglia and play a role in cerebral haemorrhage, brain trauma and other neurological diseases. Ampicillin has been reported to dose-dependently protect neurons against ischaemic brain injury, to attenuate the activity of matrix metalloproteinases and to reduce immunoreactivities of astrocytes and microglia by increasing the level of glutamate transporter-1.39 Ciprofloxacin also was claimed to inhibit the activation of the TLR4/NF-κB signalling pathway.40 Thus, GF mice may have been a more ideal model in which to conduct these experiments. Second, the translational value of the present data is limited, since experimental animals live in a very controlled environment and are fed with a standard diet, whereas the human microbiome is influenced by a huge range of factors.
Data availability statement
Data are available upon reasonable request. The data contains the original images as well as the original files, the raw data are available from the authors CL and CZ (leo_liuchang@163.com and cszhao@cmu.edu.cn). Permission to reuse the data need to be granted with the permission of the journal and the corresponding author.
Ethics statements
Patient consent for publication
Ethics approval
The study protocol was approved by the Institutional Animal Care and Use Committee of China Medical University (permit number: SCXK (Liao) 2013–0007).
References
Supplementary material
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors XL performed immunofluorescence staining. FL performed proof assistance. ZL and YZ performed behavioural tests. CL and MG performed data analysis. XC, SZ and TX provided expert technical assistance. CL and CZ conceived and designed experiments. CL and JJ wrote and edited the manuscript. CZ corrected and approved the final version of the manuscript, and is responsible fo the overall content.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.