Article Text

Abstract
Background Postsynaptic density protein-95 (PSD95) plays an important role in cerebral ischaemia injury, but its mechanism needs further research. This study aimed to explore the role of PSD95 in (Ang-(1-7))-Mas-mediated cerebral ischaemia protection and its regulatory mechanism.
Methods Oxygen–glucose deprivation (OGD) neuron and rat middle cerebral artery occlusion (MCAO) models were used as in vitro and in vivo models, respectively. TAT-MAS9C was used to disrupt the interaction between PSD95 and Mas. The recombinant PSD95 adenovirus (Ad-PSD95) was used to overexpress PSD95 in neurons.
Results Results showed that in OGD neurons, Ang-(1-7) could promote cell viability; reduce cell apoptosis; reduce the cell membrane localisation of Mas; upregulate the expression levels of pAKT, bcl-2 and I-κB; and downregulate the expression levels of Bax, pI-κB, tumour necrosis factor alpha and interleukin-1β. TAT-MAS9C could enhance the aforementioned effects of Ang-(1-7). However, the PSD95 overexpression inhibited the aforementioned effects of Ang-(1-7). In the MCAO rat model, the 2,3,5-triphenyltetrazolium chloride (TTC) staining showed that Ang-(1-7) reduced the infarct volume. The Morris water maze test showed that the number of crossings over the platform area in the Ang-(1-7) group was significantly increased. TAT-MAS9C could promote the protective effect of Ang-(1-7).
Conclusions Results suggested that PSD95 alleviated the activation of AKT and the inhibition of nuclear factor kappa B signalling pathway mediated by the Ang-(1-7)–Mas complex, thereby reducing neuronal activity, increasing apoptosis and inhibiting the Ang-(1-7)–Mas-mediated cerebral ischaemia protection.
- brain
- stroke
- cerebral infarction
Data availability statement
Data are available upon reasonable request. The raw data will be made available by the authors upon reasonable request.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
WHAT IS ALREADY KNOWN ON THIS TOPIC
Angiotensin-(1-7) (Ang-(1-7)) plays a protective role in cerebral ischaemia through Mas receptor. Our previous study found that postsynaptic density protein-95 (PSD95) has a strong interaction with Mas. However, whether PSD95 could regulate Ang-(1-7)–Mas-induced cerebral ischaemia protection and its mechanism was unknown.
WHAT THIS STUDY ADDS
This study showed that PSD95 could inhibit the Ang-(1-7)–Mas-induced cerebral ischaemia protection by reducing theinternalisation of the Mas receptor.
HOW THIS STUDY MIGHT AFFECT RESEARCH, PRACTICE AND/OR POLICY
This study will provide a new theoretical basis for the drug development of prevention and treatment of cerebral ischaemia-related diseases.
Introduction
Cerebral ischaemia is one of the most important causes of human death and disability.1 The cerebral blood flow interruption and subsequent blood flow recovery can cause a series of rapid cascade reactions, including energy metabolism disorders, excitatory amino acid release, oxidative stress, intracellular calcium homeostasis imbalance, mitochondrial dysfunction and apoptosis pathway activation, in the brain tissue and eventually lead to the death of nerve cells in the ischaemic area.2 3 The recombinant tissue plasminogen activator can restore blood flow through thrombolysis and has good therapeutic effect on acute ischaemic stroke. However, thrombolytic therapy must be administered within 4.5 hours of stroke onset. Therefore, only a small number of patients with stroke can truly undergo thrombolytic therapy, which has many limiting factors (such as low vascular recanalisation rate and possible bleeding risk).4 New drugs should be developed for the prevention and treatment of cerebral ischaemia.
Cerebral ischaemia disease is closely related to the renin–angiotensin system (RAS). RAS is an important system for regulating hydroelectrolyte balance and blood pressure.5 RAS contains two axes, that is, ACE-angiotensin II-angiotensin type 1 receptor and ACE2-angiotensin-(1-7) (Ang-(1-7))-Mas axes, which show opposite effects.6 7 Ang-(1-7) and Mas are abundant in the nervous system.8 9 Recent studies found that the ACE2–Ang-(1-7)–Mas axis has a protective effect on the cerebral ischaemia disease.5 Through the Mas receptor, Ang-(1-7) can inhibit the activity of NF-κB, reduce the expression of its downstream proinflammatory genes (including tumour necrosis factor alpha (TNF-α) and interleukin (IL)-1β), reduce inflammation and play a protective role in cerebral ischaemia.10 11 However, current studies on the protective effect of ACE2–Ang-(1-7)–Mas axis are focused on ischaemic penumbra and microglial cells. Reports on the protective effect of this axis on ischaemic neurons in vitro are few. Inflammation is the main cause of ischaemic neuronal apoptosis.12 13 Therefore, the regulatory effect of the ACE2–Ang-(1-7)–Mas axis on the inflammatory pathway in neurons remains to be further studied.
Mas is a G protein-coupled receptor with seven transmembrane structures. In Mas-transfected 293 T cells, the binding of Ang-(1-7) to Mas causes the internalisation of the Ang-(1-7)–Mas complex through endocytic vesicles. The fusion of vesicles with early endosomes activates downstream AKT and ERK pathways.14 Considering that the internalisation of Mas receptors activates AKT and ERK, the regulation of the Mas cell membrane localisation directly affects the Ang-(1-7)–Mas-induced AKT and ERK pathways. The AKT pathway is an important signalling pathway that promotes cell survival. Studies found that the activation of the AKT pathway can play a protective role in cerebral ischaemia injury by inhibiting apoptosis.15–17 However, the role of Ang-(1-7)–Mas-induced AKT pathway in cerebral ischaemia protection remains to be further studied.
Postsynaptic density protein-95 (PSD95) is a scaffolding protein with three PSD-95/Discs large/Zonula occludens-1 (PDZ) domains that are critical for the assembly of excitatory synaptic signalling complexes.18 PSD95 is a member of the membrane-associated guanylate kinase family.19 PSD95 is found to be involved in N-methyl-D-aspartic acid receptor (NMDAR)-mediated cerebral ischaemia injury. During cerebral ischaemia, the interaction between PSD95 and NMDAR is strengthened, and a large number of Ca2+ flow through NMDAR. Mediated by PSD95, the downstream signalling pathway of NMDAR is activated, leading to the death of neurons and cerebral ischaemia injury.20 21 This phenomenon suggests that PSD95 plays an important role in cerebral ischaemia injury. However, its mechanism needs to be further elucidated.
Our previous study found that PSD95 has a strong interaction with Mas. When Mas and PSD95 plasmids are cotransfected into BHK cells, they are colocalised in the plasma membrane.22 We hypothesise that PSD95 may regulate Ang-(1-7)–Mas-induced cerebral ischaemia protection through its influence on Mas localisation in the plasma membrane. In the present study, the in vivo model of middle cerebral artery occlusion (MCAO) and the in vitro model of oxygen–glucose deprivation (OGD) neurons are used to explore the effect of PSD95 on the protection of Ang-(1-7)–Mas-induced cerebral ischaemia and its regulatory mechanism. This study will explore the new mechanism of PSD95 that regulates cerebral ischaemia injury and provide a new theoretical basis for the drug development of prevention and treatment of cerebral ischaemia-related diseases.
Materials and methods
Primary neurons culture and OGD model
Primary cortical neurons were isolated from embryonic days 17–19 pregnant SD rat embryos. Cortical neurons were cultured in a neurobasal medium (Gibco, USA) containing L-glutamine and B27 (Gibco). Neurons were kept in a CO2 thermostatic incubator at 37°C and 5% CO2 humidified atmosphere. Seven-day-old cultures were adopted for all experiments.
For OGD, cells were switched to glucose-free DMEM (Gibco) and in a modular incubator chamber (Billups Rothenberg, Del Mar, California, USA) containing a mixture of 95% N2 and 5% CO2 for 1 hour. Then, cells were switched to neurobasal medium and returned to the CO2 incubator for reoxygenation for 24 hours. Ang-(1-7) (10−6 M) or A779 (10−5 M) was administered from OGD until the end of 24 hours of reoxygenation. The experiment was divided into five groups, that is, negative control, OGD, Ang-(1-7), Ang-(1-7)+A779 and Ang-(1-7)+TAT-MAS9C groups.
Cell culture
The mouse neural stem cell line NE-4C was obtained from Shanghai Zhongqiaoxinzhou Biotech Co. (Shanghai, China) and cultured in special cell culture medium for NE-4C cells (Procell Life Science & Technology Co., Wuhan, China). Cells were cultured in a 5% CO2 cell incubator and passaged every 2 days.
Design and synthesis of the penetrating membrane peptide TAT-MAS9C
TAT-MAS9C was designed on the basis of the C-terminal sequence of the Mas protein interacting with PSD95. TAT-MAS9C consisted of 9 amino acids in the carboxyl terminal of Mas and 11 amino acids in the transmembrane domain of the HIV-1 Tat protein. TAT-MAS9C was synthesised by Shanghai Bootech BioScience & Technology Co. (Shanghai, China) and labelled with rhodamine to observe the effect of membrane penetration.
Adenovirus infection
The recombinant PSD95 adenovirus (Ad-PSD95) was provided by Shanghai Genechem Co. (Shanghai, China). Neurons were cultured in a 6-well or 96-well plate for about 7 days and infected with Ad-PSD95 or Ad-control at multiplicity of infection (MOI) of 100.
Interference effect of TAT-MAS9C as detected by coimmunoprecipitation
Flag-Mas and PSD95 were transfected simultaneously into 293 cells. The experiment was divided into three groups. After transfection for 24 hours, 293 cells were incubated with or without TAT-MAS9C for 1 hour. Cells were lysed at 4°C for 1 hour, and the supernatant was collected by centrifugation and added with anti-Flag M2 Affinity Gel (Sigma-Aldrich, St. Louis, Missouri, USA). The mixture was mixed at 4°C for 3 hours. After centrifugation, the precipitate was collected and washed. The loading buffer was added to the precipitated complex and kept at 95°C for 5 min. After centrifugation, the supernatant was collected for western blot.
Cell counting Kit-8 (CCK8) cell viability assay
The CCK8 (Solarbio Biotechnology, Beijing, China) was used to assess the viability of neurons. Primary neuron cultures (4×104 cells per well) were cultured in a 96-well plate and incubated for 7 days. After OGD/reoxygenation, the CCK8 solution was added into the cells for 2 hours at 37°C. The absorbance values at 450 nm were measured using a microplate reader. The relative cell viability was quantified using the absorbance of each group normalised to that of the control group.
Detection of the MAS protein localisation by flow cytometry
NE-4C cells were cultured in six-well plates. When the cell density reached 80%, cells were treated with OGD in Hank's balanced salt solution (HBSS) for 1 hour at 37°C, stimulated for 25 min with 10−5 M TAT-MAS9C or/and 10−6 M Ang-(1-7) in HBSS at 37°C, washed with cold HBSS and treated with anti-Mas antibody (1:200; Novus Biologicals, Littleton, Colorado, USA) in HBSS for 50 min on ice. Cells were then washed with cold HBSS salt solution and treated with Alexa Fluor 488 goat anti-rabbit IgG (1:1000) for 50 min on ice. Then, NE-4C cells were washed with cold HBSS salt solution and digested with 5 mM EDTA in phosphate buffered saline (PBS on ice. The suspended NE-4C cells were collected by centrifugation and fixed with 4% paraformaldehyde. Samples were analysed using a flow cytometer (BD Biosciences, Mississauga, Ontario).
Detection of the MAS protein localisation by immunoprecipitation
Neurons were cultured in 60 mm dishs and treated with OGD in HBSS for 1 hour at 37°C. Cells were then stimulated with Ang-(1-7) (10−6 M) or Ang-(1-7) (10−6 M)+TAT-MAS9C (10−5 M) in HBSS at 37°C for 25 min, washed with cold HBSS, and treated with anti-Mas antibody (1:100; Santa Cruz Biotechnology, Santa Cruz, California, USA) in HBSS for 50 min on ice. Cells were lysed, and the supernatant was collected by centrifugation. Protein A/G was added into the supernatant and incubated overnight. After centrifugation, the precipitate was collected and washed. The precipitated complex was resuspended in protein loading buffer, heated at 95°C for 5 min and centrifuged. The supernatant was used for western blot.
Western blot
Protein samples were run on sodium dodecyl sulfate polyacrylamide gels and transferred onto the polyvinylidene difluoride (PVDF) membrane. Membranes were blocked in 5% milk for 1 hour and then incubated overnight with anti-p-IκB-α, anti-IκB-α, anti-pAKT, anti-Bcl-2, anti-Bax, anticleaved caspase-3 (Cell Signaling Technology, Beverly, Massachusetts, USA), anti-FLAG (Sigma-Aldrich), anti-PSD95 (Abcam, Cambridge, Massachusetts, USA), anti-Mas (Novus Biologicals) and anti-β-actin (ZSGB-BIO, Beijing, China). After washing with PBS three times, the membrane was incubated with horseradish peroxidase-conjugated secondary antibodies (Cell Signaling Technology) for 1 hour. Protein bands were detected by enhanced chemiluminescence detection reagents (Applygen Technologies, Beijing, China).
Reverse-transcription quantitative real-time PCR
Neurons were treated and suspended in the RNAiso Plus (Takara, Otsu, Japan). Total RNA was extracted and converted into cDNA by using the PrimeScript RT reagent kit (Takara). The cDNA was amplified by quantitative PCR (qPCR) through the SYBR Premix Ex Taq II (Takara). The primers used were as follows: TNF-α forward primer, GCCACCACGCTCTTCTGTCTAC; TNF-α reverse primer, GGGTCTGGGCCATAGAACTGAT; IL-1β forward primer, ACCTTCCAGGATGAGGACATGA; and IL-1β reverse primer, CTAATGGGAACGTCACACACCA. The relative mRNA expression levels of TNF-α and IL-1β normalised to β-actin were analysed using the 2−ΔΔCt method.
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labelling (TUNEL)
Neurons were cultured on coverslips placed in 24-well plates. The TUNEL Assay Kit (Roche, Basel, Switzerland) was used to assess the apoptosis of neurons. The cultured neurons were fixed with 4% paraformaldehyde, permeated with 0.1% Triton X-100, incubated with TUNEL solution for 1 hour at 37°C and washed with phosphate-buffered saline with Tween 20 (PBST). The coverslips were mounted and samples were visualised under a fluorescence microscope. The percentage of positive cells in the total neurons was used to calculate the apoptotic rate.
Animals and treatment
Male, SPF-grade (Shanghai SLAC Laboratory Animal Co., certificate number 20170005030024) SD rats (6–8 weeks old, 180–220 g) were selected and divided into four groups, that is, sham control, MCAO, Ang-(1-7) and TAT-MAS9C+Ang-(1-7) groups. Animals were randomly grouped indiscriminately by means of a table of random numbers. There were six rats in each group, a total of 48 rats. Six animals died after MCAO.
Middle cerebral artery occlusion
After anaesthesia, the right common carotid artery (CCA), internal carotid artery (ICA) and external carotid artery (ECA) were separated, and CCA and ECA were ligated. A small incision was cut near the branch of CCA, and a filament was inserted into ICA through ECA to block the blood supply of the cerebral artery. The filament was removed 90 min after ischaemia to produce reperfusion for 30 hours. The artificial cerebrospinal fluid, Ang-(1-7) (2.5 nmol) or Ang-(1-7) (2.5 nmol)+TAT-MAS9C (25 nmol) was injected slowly into the lateral ventricle 15 min after MCAO once a day.
Neurological deficit score
The neurological performance of rats was graded using Longa’s test: 0, no neurological function deficit; 1, flexion of the left forelimb when the tail was lifted; 2, reclining to the left when crawling; 3, reclining to the left at rest; and 4, disorders of consciousness or death. A score of 1–3 could be considered as successful modelling of MCAO rats. All data are recorded in a double-blinded manner.
2,3,5-Triphenyl tetrazolium chloride (TTC) staining
The brain tissue was frozen at 4°C for 10 min, and coronal sections were collected. Sections were stained with 2% TTC solution and incubated at 37°C for 30 min in the dark. The normal tissue remained red, whereas the infarct area turned white. The ratio of the infarct area to the total brain area was calculated using ImageJ software.
Morris water maze test
The Morris water maze was used to observe the spatial learning and memory ability of MCAO rats. For cued training, rats successively entered the water from four quadrants facing the pond wall. If no platform was found within 90 s, rats were guided to the platform and rested on the platform for 30 s. Cued training was done three times a day for 5 days. On day 6, the platform was removed from the pool, and each rat underwent a 90 s swimming probe test. Swimming tracks and the number of crossings over the platform area were monitored using the Morris water maze video analysis system. The test was recorded in a double-blinded manner.
Statistical analysis
Experimental results were processed using GraphPad Prism V.7.0 software. Data were presented as mean±SD. Student’s t test was used for comparison between the two groups, and one-way analysis of variance was used for comparisons among three or more groups. A p value of <0.05 indicated that the difference was statistically significant.
Results
TAT-MAS9C destroys the interaction between PSD95 and Mas
TAT-MAS9C was used as the main tool to study the regulatory effect of PSD95 on Mas. TAT-MAS9C was designed on the basis of the C-terminal sequence of the Mas protein interacting with PSD95. TAT-MAS9C was speculated to bind PSD95, thereby destroying the binding between Mas and PSD95 (figure 1A). In addition, TAT-MAS9C contained 11 amino acids in the transmembrane domain of the HIV-1 Tat protein, which was theoretically speculated to be able to penetrate the cell membrane. Neurons were incubated with rhodamine-labelled TAT-MAS9C for 0, 10, 30 and 90 min. As shown in figure 1B, the entrance of TAT-MAS9C in the cells increased in a time-dependent manner. After 30 min of incubation, the concentration of TAT-MAS9C entering the cells reached the maximum, whereas the negative control could hardly enter the cells (data not shown).

TAT-MAS9C destroys the interaction between PSD95 and Mas. (A) Hypothesis: the Mas–PSD95 complex may be disrupted by TAT-MAS9C. (B) The rhodamine-labelled TAT-MAS9C (10−5 M) accumulation in neurons was visualised after incubation for 0, 10, 30 and 90 min. (C) PSD95 was subjected to coimmunoprecipitation with Flag-Mas in HEK293 cells treated with 10−5 M TAT-MAS9C. (D) PSD95 was subjected to coimmunoprecipitation with NR2B in 293 cells treated with 10−5 M TAT-MAS9C. PSD95, postsynaptic density protein-95.
The effect of TAT-MAS9C on the interaction between Mas and PSD95 was detected by coimmunoprecipitation assays. As expected, 10−5 M TAT-MAS9C incubation detected less PSD95 content in the immunoprecipitation complex (figure 1C), indicating that TAT-MAS9C could destroy the interaction between PSD95 and Mas. HEK293 cells were transfected with NR2B and PSD95 plasmids, which were closely related to cerebral ischaemia, to confirm that TAT-MAS9C could not destroy the binding of other proteins to PSD95, and coimmunoprecipitation was performed. Results showed that TAT-MAS9C could not destroy the interaction between NR2B and PSD95 (figure 1D). Therefore, the specificity of TAT-MAS9C was further confirmed.
Effects of PSD95 on Ang-(1-7)–Mas-induced cerebral ischaemia protection in OGD neurons
Neurons were cultured in 6-well or 96-well plate for about 7 days and subjected to TAT-MAS9C intervention or Ad-PSD95 infection. Cell viability was detected using the CCK8 assay. As shown in figure 2A, the neuron viability in the OGD group was significantly lower than that in the control group, indicating that the OGD model was successful (p<0.05). Compared with the OGD group, the Ang-(1-7) group had significantly increased neuronal viability. Compared with the Ang-(1-7) group, the Ang-(1-7)+A779group had significantly decreased neuronal viability. Given that A779 was a specific inhibitor of Mas receptor, the protective effect of Ang-(1-7) on OGD neurons was suggested to be realised through the Mas receptor. The effect of TAT-MAS9C on the viability of Ang-(1-7)-treated neurons was then detected. Compared with that in the Ang-(1-7) group, the viability of neurons in the Ang-(1-7)+TAT-MAS9C group was significantly increased, indicating that the destruction of the interaction between Mas and PSD95 increased the cell viability. Neurons were infected with Ad-PSD95 to verify the function of PSD95 on the viability of Ang-(1-7)-treated neurons, and the overexpression of PSD95 was confirmed by western blot (figure 2B). PSD95 overexpression significantly reduced the viability of neurons treated with Ang-(1-7) (figure 2C). These results suggested that PSD95 reduced the viability of neurons treated with Ang-(1-7) through its interaction with Mas.

Effects of PSD95 on the Ang-(1-7)–Mas-induced cerebral ischaemia protection in OGD neurons. Neurons were incubated with Ang-(1-7) (10−6 M), A779 (10−5 M) or TAT-MAS9C (10−5 M) during OGD. After OGD for 1 hour, cells were switched to normal condition for 24 hours. For Ad-PSD95 infection, neurons were infected with Ad-PSD95 or Ad-control at MOI of 100 for 3 days. Then, neurons were incubated with Ang-(1-7) (10−6 M) for 1 hour of OGD treatment and returned to normal conditions for 24 hours. (A) The effect of TAT-MAS9C on cell viability was measured using the CCK8 assay in Ang-(1-7)-treated OGD neurons. The data within each group were normalised to those of the control (n=6; vs control group*, vs OGD group&, vs Ang-(1-7) group#). (B) The overexpression of PSD95 by Ad-PSD95 infection was detected by western blot. (C) The effect of Ad-PSD95 on cell viability was measured using the CCK8 assay in Ang-(1-7)-treated OGD neurons (n=6, vs Ad-control*, vs OGD group&, vs Ad-control+Ang-(1-7)# group). (D) The effect of TAT-MAS9C on the apoptosis of neurons was identified by TUNEL staining and quantified using the percentage of TUNEL-positive cells. The data within each group were normalised to those in the control group. (E) The effect of PSD95 overexpression on the apoptosis of neurons was identified by TUNEL staining and quantified using the percentage of TUNEL-positive cells. The data within each group were normalised to those in the Ad-control group. *P<0.05, &P<0.05, #P<0.05. Ad-PSD95, PSD95 adenovirus; Ang-(1-7), angiotensin-(1-7); OGD, oxygen–glucose deprivation; PSD95, postsynaptic density protein-95; TUNEL, terminal deoxynucleotidyl transferase-mediated dUTP nick-end labelling; DAPI, 4',6-diamidino-2-phenylindole.
Cell viability is closely related to apoptosis. Cerebral ischaemia can induce apoptosis. TUNEL staining was then used to detect cell apoptosis. As shown in figure 2D, compared with the OGD group, the control and Ang-(1-7) groups had significantly lower proportions of TUNEL-positive neurons. Compared with that of the Ang-(1-7) group, the proportions of TUNEL-positive neurons of the Ang-(1-7)+A779 and Ang-(1-7)+TAT-MAS9C groups were significantly increased and significantly decreased, respectively. The Ad-PSD95 infection was performed to verify the function of PSD95 on the apoptosis of Ang-(1-7)-treated neurons. As shown in figure 2E, PSD95 overexpression significantly increased the proportion of TUNEL-positive neurons. These results suggested that PSD95 increased the apoptosis of neurons treated with Ang-(1-7) through its interaction with Mas.
Effect of PSD95 on the plasma membrane localisation of the Mas protein in OGD neurons treated with Ang-(1-7)
Mas was reported to be retained in the plasma membrane by interacting with PSD95 in BHK cells22 . Ang-(1-7) could promote the internalisation of Mas receptors. Next, we used TAT-MAS9C to investigate whether PSD95 could affect the localisation of Mas protein in the plasma membrane of Ang-(1-7)-treated OGD neurons. Protein immunoprecipitation was used to detect the expression level of Mas in the plasma membrane. Compared with Ang-(1-7) treatment alone, the TAT-MAS9C and Ang-(1-7) cotreatment significantly reduced the Mas content in the immunoprecipitation complex (figure 3A). The Mas receptor on the cell membrane of OGD NE-4C cell was labelled with the Mas antibody, and the effect of TAT-MAS9C on the plasma membrane localisation of Mas was detected by flow cytometry. As shown in figure 3B,C, the fluorescence intensity on the surface of NE-4C cells in the TAT-MAS9C treatment group was significantly reduced, indicating that TAT-MAS9C treatment reduced the plasma membrane localisation of Mas and promoted the internalisation of the Ang-(1-7)–Mas complex. These results suggested that PSD95 disrupted the protective effect of Ang-(1-7) on OGD neurons by reducing the internalisation of the Mas receptor.

Effect of TAT-MAS9C on the cell membrane localisation of Mas protein in OGD neurons treated with Ang-(1-7). Neurons or NE-4C cells were treated with OGD for 1 hour and then stimulated for 25 min with 10−6 M Ang-(1-7) or 10−5 M TAT-MAS9C+10−6 M Ang-(1-7) in HBSS. After washing with cold HBSS, cells were treated with anti-Mas antibody for 50 min on ice. (A) Protein immunoprecipitation was used to detect the expression level of MAS in the plasma membrane of neurons. Cell lysates were subjected to immunoprecipitation with protein A/G. The expression levels of MAS and PSD95 in the immunoprecipitation complex was detected by western blot. Images were representative of three independent experiments. (B) Flow cytometry was used to detect the expression level of MAS in the plasma membrane of NE-4C cells. Cells were treated with Alexa Fluor 488 goat anti-rabbit IgG for 50 min on ice, washed with cold HBSS, and digested with 5 mM EDTA in PBS on ice. Suspended NE-4C cells were collected and detected by flow cytometry. (C) The relative fluorescence corresponding to MAS on the membrane was analysed by flow cytometry. Values were obtained from three separate experiments (n=3). Ang-(1-7), angiotensin-(1-7); IB, immunoblot; IP, immunoprecipitation; OGD, oxygen–glucose deprivation; PSD95, postsynaptic density protein-95.
Effect of PSD95 on the signalling pathway in Ang-(1-7)-treated neurons
The activation of the NF-κB pathway is an important pathological mechanism leading to cerebral ischaemia injury.11 The AKT signalling pathway is a pathway that has been found to be directly activated by Mas receptor internalisation, which can promote cell survival and inhibit cell apoptosis.14 Here, the expression levels of NF-κB and AKT signalling pathway-related proteins were detected by Western blot and qPCR. Compared with the OGD group, the control and Ang-(1-7) groups had lower expression levels of p-IκB-α, Bax, TNF-α and IL-1β. Compared with those in the Ang-(1-7) group, the expression levels of p-IκB-α, Bax, TNF-α and IL-1β were increased in the Ang-(1-7)+A779 group and decreased in the TAT-MAS9C+Ang-(1-7) group. The expression levels of IκB-α, p-AKT and Bcl-2 showed an opposite trend to the expression levels of p-IκB-α, Bax, TNF-α and IL-1β (figure 4A–C). The Ad-PSD95 infection was performed to verify the function of PSD95 on the signalling pathway in Ang-(1-7)-treated neurons. As shown in figure 4D–F, PSD95 overexpression significantly increased the expression levels of p-IκB-α, Bax, TNF-α and IL-1β and decreased the expression levels of IκB-α, p-AKT and Bcl-2. These results suggested that PSD95 activated the NF-κB pathway and weakened the AKT pathway in the OGD neurons treated with Ang-(1-7).
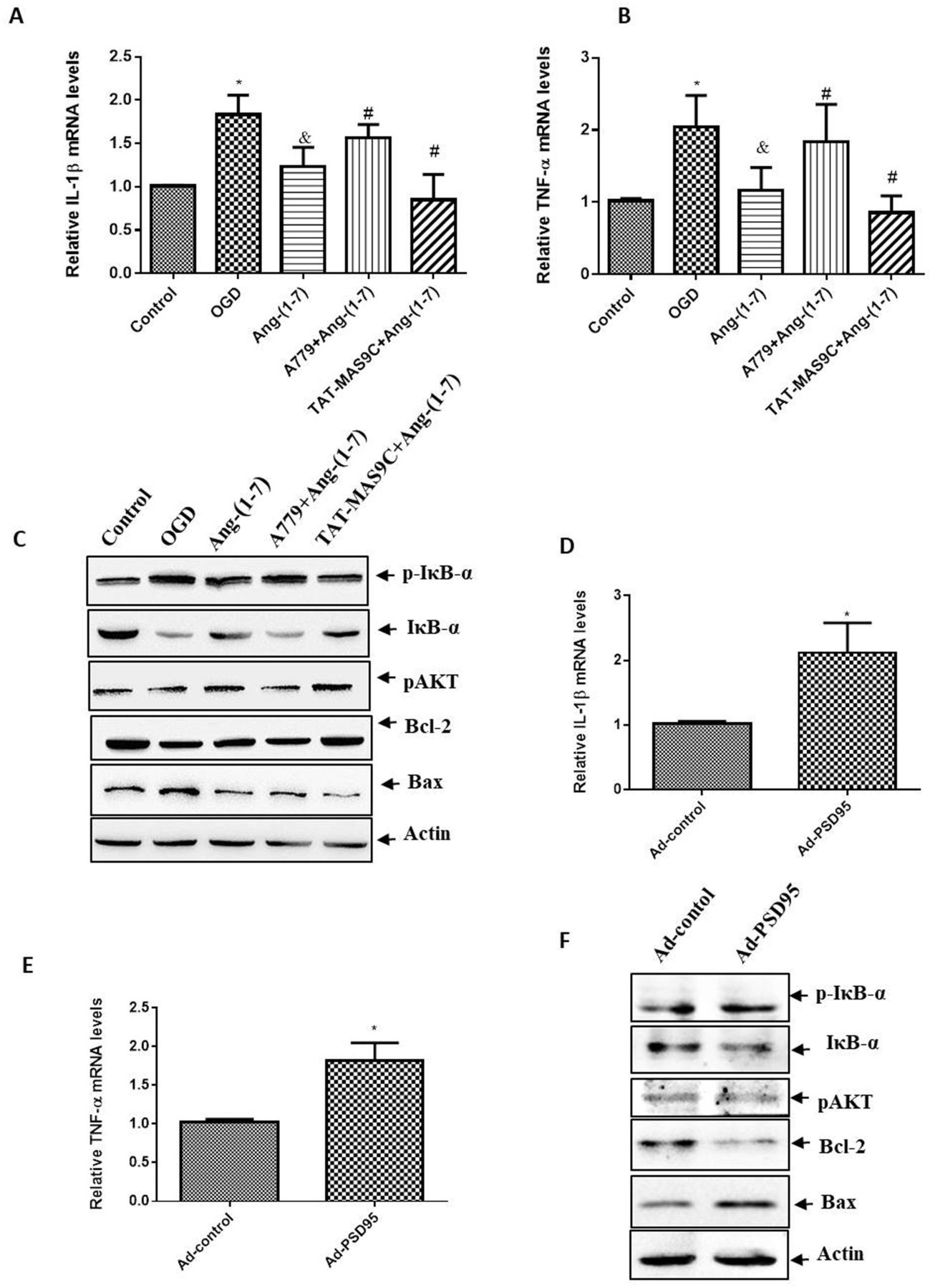
Effect of PSD95 on the signalling pathway in Ang-(1-7)-treated neurons. Neurons were incubated with Ang-(1-7) (10−6 M), A779 (10−5 M) or TAT-MAS9C (10−5 M) during OGD. After OGD for 1 hour, cells were switched to normal conditions for 24 hours. qPCR was used to analyse the expression levels of (A) IL-1β and (B) TNF-α. Values were expressed as mean±SD (n=3; vs control group*, vs OGD group&, vs Ang-(1-7) group#). (C) Western blot was used to analyse the expression levels of p-IκB-α, IκB-α, p-AKT, Bcl-2 and Bax. Images were representative of three independent experiments. For Ad-PSD95 infection, neurons were infected with Ad-PSD95 or Ad-control at MOI of 100 for 3 days. Then, neurons were incubated with Ang-(1-7) (10−6 M) for 1 hour of OGD treatment and returned to normal conditions for 24 hours. qPCR was used to analyse the expression levels of (D) IL-1β and (E) TNF-α. Values were expressed as mean±SD (n=3, vs Ad-control*). (F) Western blot was used to analyse the expression levels of p-IκB-α, IκB-α, p-AKT, Bcl-2 and Bax. Images were representative of three independent experiments. *P<0.05, &P<0.05, #P<0.05. Ad-PSD95, PSD95 adenovirus; Ang-(1-7), angiotensin-(1-7); IL, interleukin; OGD, oxygen–glucose deprivation; PSD95, postsynaptic density protein-95; qPCR, quantitative PCR; TNF-α, tumour necrosis factor alpha.
Effects of PSD95 on Ang-(1-7)–Mas-induced cerebral ischaemia protection in MCAO rats
Whether the use of TAT-MAS9C during stroke could reduce ischaemic damage in vivo was investigated. Rats were subjected to MCAO for 90 min. Ang-(1-7), TAT-MAS9C or artificial cerebrospinal fluid was injected slowly into the lateral ventricle 15 min after MCAO. The neurological deficit score was determined 30 hours after MCAO. As shown in figure 5A, the neurological deficit scores of sham, MCAO, Ang-(1-7) and TAT-MAS9C+Ang-(1-7) groups were 0, 2.5, 1.7 and 1.0, respectively. The neurological score in the TAT-MAS9C+Ang-(1-7) group was significantly improved compared with that in the Ang-(1-7) group. Furthermore, TTC staining was performed to evaluate the infarct volume 30 hours after MCAO. As shown in figure 5B,C, compared with the normal group, the MCAO group had increased infarct volume. Compared with the Ang-(1-7) group, the MCAO group had increased infarct volume, whereas the TAT-MAS9C+Ang-(1-7) group had decreased infarct volume. Then, western blot was used to detect apoptosis-related proteins in the ischaemic penumbra of cerebral cortex. Results also showed that compared with the Ang-(1-7) group, the MCAO group had increased expression of Bax and cleaved caspase-3 and decreased expression of Bcl-2, whereas the TAT-MAS9C+Ang-(1-7) group had decreased expression of Bax and cleaved caspase-3 and increased expression of Bcl-2 (figure 5D). The Morris water maze test was performed to assess spatial memory. As shown in figure 5E,F, the number of crossings over the platform area in the MCAO group was significantly reduced compared with that in the control group (p < 0.05). By contrast, compared with the Ang-(1-7) group, the MCAO group had decreased number of crossings over the platform area (p < 0.05), whereas the TAT-MAS9C+Ang-(1-7) group had an increased number of crossings over the platform area. These results showed that TAT-MAS9C could reduce neuronal apoptosis and promote Ang-(1-7)–Mas-induced cerebral ischaemia protection, suggesting that PSD95 could inhibit the cerebral ischaemia protection induced by the Ang-(1-7)–Mas complex in the MCAO model rat.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of PSD95 on Ang-(1-7)–Mas-induced cerebral ischaemia protection in MCAO rats. Rats were pretreated with artificial cerebrospinal fluid, Ang-(1-7) or Ang-(1-7)+TAT-MAS9C for 7 days prior to MCAO. Rats were then subjected to MCAO for 90 min. Ang-(1-7), A779 or TAT-MAS9C was injected slowly into the lateral ventricle 15 min after MCAO once a day. (A) The neurological performance of rats was graded using Longa’s test 30 hours after MCAO. (B) The infarct volume was evaluated 30 hours after MCAO. Brain tissue sections were stained with TTC. (C) The infarct volume was calculated as the ratio of the infarct area to the total area by using ImageJ software. (D) Western blot was used to analyse the expression levels of Bcl-2, Bax and cleaved caspase 3 in the ischaemic penumbra of the cerebral cortex. (E)The Morris water maze test was performed to assess spatial memory. Representative images are swimming tracks. (F) The number of crossings over the platform area in the Morris water maze test was analysed. Values were expressed as mean±SD (n=6, vs sham group*, vs MCAO group&, vs Ang-(1-7) group#). *P<0.05, &P<0.05, #P<0.05. Ang-(1-7), angiotensin-(1-7); MCAO, middle cerebral artery occlusion; PSD95, postsynaptic density protein-95; TTC, 2,3,5-triphenyltetrazolium chloride.
Discussion
PSD95, an important scaffolding protein at the postsynaptic density of glutamatergic excitatory synapses,23 plays an important role in the regulation of cerebral ischaemia injury24–26 and binds to the C-terminus of Mas.22 Ang-(1-7) plays a protective role in cerebral ischaemia by acting on Mas receptors.5 27 28 However, whether PSD95 regulates the protection of cerebral ischaemia induced by Ang-(1-7)–Mas remains unclear. In the present study, we demonstrated the disruptive role of PSD95 against Ang-(1-7)–Mas-induced cerebral ischaemia protection in OGD neurons via TAT-MAS9C incubation and Ad-PSD95 transduction and in MCAO rat via TAT-MAS9C injection. Several findings have been identified in our study. First, Ang-(1-7) treatment increased the viability and reduced the apoptosis of OGD neurons. The effect of Ang-(1-7) was reversed by Mas receptor antagonist A779, enhanced by TAT-MAS9C and attenuated by Ad-PSD95 transduction. Second, compared with Ang-(1-7) alone, the administration of TAT-MAS9C and Ang-(1-7) reduced neurological deficit score, infarct volume, apoptosis-related protein expression and increased the number of crossings over platform areas in MCAO rat. In addition, TAT-MAS9C incubation promoted the internalisation of Mas receptors induced by Ang-(1-7), thereby promoting the Ang-(1-7) -Mas complex-mediated signalling pathway.
PSD-95 is a potential target for protecting the brain from ischaemic injury. Studies have shown that Tat peptides were used to treat stroke by disrupting the interaction between PSD95 and its receptor. TAT peptide can be used as a transfection vector for the protein.29 Tat-NR2B9c, a transmembrane peptide containing nine amino acids in the carboxyl terminal of NMDAR and 11 amino acids in the transmembrane domain of the HIV-1 Tat protein, played a protective role in NMDAR-mediated cerebral ischaemia injury by interfering with the interaction between PSD95 and NMDAR receptor.30 In addition, Tat-GluR6-9c inhibited GluR6-mediated signalling pathway by disrupting the interaction between PS595 and GluR6 and protected neurons against ischaemic injury.31 In our present study, we synthesised the TAT-MAS9C fusion protein and delivered it to primary cortical neurons. TAT-MAS9C was speculated to inhibit the binding of PSD95 to Mas without affecting the function of PSD95. As expected, results showed that TAT-MAS9C penetrated neurons easily in a time-dependent manner and could decrease the interaction between PSD95 and Mas. Therefore, TAT-MAS9C was used as the main tool to study the regulatory effect of PSD95 on Mas. We also found that TAT-MAS9C promoted Ang-(1-7)–Mas-mediated protection against cerebral ischaemia.
The Ang-(1-7)–Mas axis has recently received attention and is proposed as a potential therapeutic target for ischaemic stroke. Ang-(1-7) contributes to the recovery of performance in rats after stroke.32 The oral delivery of Ang-(1-7) after stroke reduces infarct volume, improves neurological function and does not cause changes in blood pressure and heart rate.33 The beneficial effects of Ang-(1-7) are reversed by the administration of A779, a Mas receptor inhibitor, suggesting that the ischaemic protection of Ang-(1-7) was realised through the Mas receptor.28 34 35 Consistent with previous reports, we also found the protective effect of Ang-(1-7) on cerebral ischaemia in MCAO rats. In addition, we found the protective effect of Ang-(1-7) on OGD neurons through Mas receptor for the first time, providing a cellular model for studying the mechanism by which PSD95 disrupted the protective effect of Ang-(1-7)–Mas on cerebral ischaemia.
PSD95 contains three PDZ domains. Through the different PDZ domains, PSD95 can interact with a variety of receptors containing PDZ-binding motifs to regulate the function of receptors.36 For example, PSD95 can regulate the potassium ion channel by binding to the KV1 receptor.37 PSD95 can inhibit the β1AR desensitisation by binding to the β1 adrenergic receptor.38 PSD95 regulated the CRFR1 localisation by interacting with CRFR1 and β-arrestin2.39 40 PSD95 and Mas were reported to colocate on the cell membrane in BHK cells.22 In the present study, immunoprecipitation and flow cytometry showed that TAT-MAS9C could promote the internalisation of Mas receptors in cells treated with Ang-(1-7), suggesting that PSD95 disrupted the protective effect of Ang-(1-7) on OGD neurons by reducing the internalisation of the Mas receptor.
Considering that PSD95 could affect the localisation of Mas in OGD neurons treated with Ang-(1-7) and that Ang-(1-7) could activate the downstream AKT and inflammatory pathways mediated by Mas, we next investigated whether PSD95 could affect the Mas-mediated AKT and inflammatory pathways in OGD neurons treated with Ang-(1-7). As expected, the TAT-MAS9C treatment and PSD95 overexpression showed that PSD95 could disrupt the activation of the AKT pathway and the inhibition of the inflammatory pathway in Ang-(1-7)-treated OGD neurons.
In summary, our experiments suggested that PSD95 could affect the cell membrane localisation of Mas, thus affecting the downstream AKT and inflammatory pathways mediated by Mas and inhibiting the protective effect of Ang-(1-7)–Mas on cerebral ischaemia injury.
Data availability statement
Data are available upon reasonable request. The raw data will be made available by the authors upon reasonable request.
Ethics statements
Patient consent for publication
Ethics approval
All animals were treated in accordance with the international guidelines for animal research, and the study design was approved by the Institutional Animal Care and Use Committee of Binzhou Medical University (PJ2018-03-27)
References
Footnotes
BZ, ZW and XL contributed equally.
Contributors WB designed the research and acted as guarantor; BZ, ZW, XL, KL and LY performed the research; XW analysed data; WB and YS wrote the manuscript; JJ, CX and DZ revised the manuscript.
Funding This study was supported by the National Natural Science Foundation of China (number 81801192) and the Project of Shandong Province Higher Educational Science and Technology Program (number J18KA141).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.